

Review Article
Decoding The Gut-Brain-Microbiome Axis In GLP-1 Signaling
H. P. Ghosh Research Center, New Town, Kolkata, West Bengal 700161, India.
*Corresponding Author: Swarup K. Chakrabarti, H. P. Ghosh Research Center, New Town, Kolkata, West Bengal 700161, India.
Citation: Chakrabarti SK. (2026). Decoding The Gut-Brain-Microbiome Axis In GLP-1 Signaling, Journal of Clinical Research and Clinical Trials, BioRes Scientia Publishers. 5(1):1-15. DOI: 10.59657/2837-7184.brs.26.066
Copyright: © 2026 Swarup K. Chakrabarti, this is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: March 11, 2026 | Accepted: March 27, 2026 | Published: April 03, 2026
Abstract
The treatment of type 2 diabetes and obesity has been reshaped by glucagon-like peptide-1 receptor agonists (GLP-1RAs), which produce substantial weight loss along with clear metabolic benefits. Increasing evidence shows that these drugs act beyond classical endocrine pathways, engaging interconnected biological systems that link the gut microbiome with the gastrointestinal tract, immune system, and brain. However, much of the existing research on GLP-1RA-induced microbiome changes remain largely associative, and the mechanisms through which these microbial shifts influence neuroendocrine regulation, adipose tissue remodeling, and preservation of lean mass are still poorly understood. This review integrates findings from preclinical and clinical studies to examine how GLP-1RAs alter gut microbial composition and how microbial metabolites shape hormone secretion, immune balance, gut–brain communication, and tissue-specific metabolic responses. Across studies, GLP-1RAs consistently promote short-chain fatty acid–producing bacteria while suppressing pro-inflammatory taxa. By framing the gut microbiome as an active partner in GLP-1-mediated effects rather than a passive bystander, this review highlights new opportunities for developing more personalized and durable metabolic therapies.
Keywords: GLP-1 receptor agonists; gut microbiome; gut-brain axis; short-chain fatty acids; adipose tissue remodeling; neuroendocrine signaling; sarcopenia
Introduction
The main therapies for type 2 diabetes (T2D) and obesity today consist of glucagon-like peptide-1 receptor agonists (GLP-1RAs), which not only effectively control blood glucose levels but also encourage considerable weight loss and reduce cardiovascular risk [1-4]. The metabolic impacts of GLP-1RAs go beyond their conventional functions, as they operate through a variety of interconnected systems linking the gastrointestinal (GI) tract, central nervous system (CNS), and immune system [5-7]. This integrated signaling network, now acknowledged as the gut-brain-microbiome axis, manages nutrient detection, appetite regulation, inflammation, and energy equilibrium, highlighting that GLP-1 acts at the convergence of metabolic and neuroendocrine control [8-10]. Treatment with GLP-1RAs during the initial year typically results in weight loss ranging from 15% to 25% [11,12]. However, the long-term sustainability of these effects remains uncertain, as many patients tend to regain weight upon discontinuation of therapy. The need for enduring and effective interventions is further underscored by the prevalence of GI side effects and rare but serious complications, such as pancreatitis and bowel obstruction, alongside the critical importance of preserving lean body mass [13,14]. While most studies indicate that fat loss is more pronounced than skeletal muscle loss, the biological mechanisms governing tissue-specific remodeling during GLP-1-induced weight loss remain insufficiently understood [15].
Importantly, the gut microbiome functions as an adaptive system that controls metabolism, immune reactions, and neuroendocrine signaling in the host [16-18]. GLP-1RAs affect microbial populations by encouraging the proliferation of bacteria that produce short-chain fatty acids (SCFAs), such as Akkermansia, Bifidobacterium, Roseburia, and Faecalibacterium, while inhibiting pro-inflammatory and unbalanced species [19-21]. Microbial byproducts, such as butyrate and propionate, strengthen the intestinal barrier, enhance insulin sensitivity, modulate appetite regulation, and decrease systemic inflammation [22,23]. These byproducts also activate intestinal L cells, boosting the secretion of endogenous GLP-1 and creating a reciprocal feedback mechanism between the host and its microbes [24,25]. Overall, these results indicate that the gut microbiome acts not just as a passive responder but as an active player in mediating the effectiveness of GLP-1.
Despite these advancement, notable gaps still exist. The majority of research is correlational, which restricts our ability to understand how alterations in the microbiome affect metabolic and neurobehavioral results. The microbial gene signatures that predict responses to GLP-1 vary inconsistently across various drug formulations and research designs, which reflects differences in treatment duration, dietary management, initial metabolic conditions, and additional medications [26,27]. Although independent relationships have been identified between GLP-1 therapy, gut microbiota composition, and skeletal muscle, direct evidence does not exist to show that microbial changes induced by GLP-1 provide protection against muscle loss during weight reduction caused by pharmacological means. This concern is especially relevant due to the potential risk of sarcopenia and loss of function during significant weight loss. 28 Similarly, the ways in which microbial signals influence brain insulin and leptin sensitivity, neuroinflammation, and the circuitry of the hypothalamus are still largely uncharted, as the integration of microbiome research with studies of the CNS has yet to be fully realized [29,30]. Additionally, our capacity to monitor temporal changes and establish causal relationships across the gut-brain-microbiome axis is further complicated by the absence of longitudinal, mechanistic, and multi-omics studies [31,32].
This review thus consolidates existing preclinical and clinical studies to investigate the relationships between GLP1-RAs, the gut microbiome, and the gut–brain axis (GBA). The first parts present findings that show how different GLP-1 analogs affect microbial diversity, composition, and metabolic function. Following this, the review discusses how alterations in the gut microbiome due to GLP-1RA influence neuroendocrine signaling, aid in correcting immune imbalances, and promote overall metabolic equilibrium. The article also explores how these effects impact the remodeling of adipose tissue (AT) and emphasizes the potential danger of muscle loss, such as sarcopenia, which has become a notable concern with GLP-1 analog therapy. By identifying the gut microbiome as an active player in the effects mediated by GLP-1 instead of a mere passive factor, this review puts forth a conceptual framework designed to clarify the variability in therapeutic responses and to inform the creation of personalized GLP-1 strategies that take the microbiome into account.
Impact of GLP-1 Analogs on Gut Microbiota: Preclinical and Clinical Insights
GLP-1RAs, such as liraglutide, semaglutide, and dulaglutide, are increasingly acknowledged for their potential to affect gut microbiota in addition to their well-known functions in regulating blood glucose levels and aiding in weight loss [1-4]. Initial studies in animals suggest that these agonists effectively contribute to maintaining microbiome equilibrium by fostering optimal conditions for the proliferation of beneficial microorganisms like Akkermansia muciniphila, Bacteroides, Lactobacillus, Roseburia, and Faecalibacterium prausnitzii [33,34]. Certain microorganisms excel at generating SCFAs, such as butyrate and propionate, which have demonstrated the ability to improve insulin sensitivity, bolster the intestinal barrier, and decrease systemic inflammation [23,24]. In terms of mechanism, GLP-1RAs may improve gut health by influencing gut motility, altering luminal pH, and changing bile acid metabolism, thereby encouraging the growth of beneficial bacteria and supporting the gut-brain-microbiome connection [7,21,22,24,25,35].
The evidence presented in Table 1 [36-66] substantiates these effects. Liraglutide significantly influences the microbiota by increasing the relative abundance of Akkermansia, Bacteroides, Lactobacillus, Parabacteroides, and Oscillospira across various rodent models [36-38]. In models following a methionine- and choline-deficient (MCD) diet, liraglutide not only restores Bacteroides populations but also alters the Erysipelotrichaceae composition from Allobaculum to Turicibacter, indicating its potential to prevent diet-induced dysbiosis in the context of liver disease [39,40]. In male db/db mice, an increase in short-chain fatty acid (SCFA)-producing bacteria (such as Parabacteroides, Oscillibacter, and Prevotellaceae) coincides with a reduction in pro-inflammatory genera (like Anaerotruncus and Lachnospiraceae) and a positive change in alanine aminotransferase/aspartate aminotransferase (ALT/AST) levels, thereby establishing a clear connection between changes in the microbiome and metabolic as well as liver health benefits [41-43].
Table 1: GLP-1 Analog Effects on Gut Microbiota.
| GLP-1 Analog | Model / Study Type | Microbiota Changes (Composition / Diversity) | Functional / Metabolic Outcomes |
| Liraglutide | Animal (rodent models) | ↑ Beneficial genera (e.g., Akkermansia, Bacteroides, Lactobacillus, Parabacteroides, Oscillospira); ↓ harmful taxa; ↓ Firmicutes:Bacteroidetes ratio in some models | Enhanced glucose metabolism, reduced weight gain, improved lipid profiles [36-38] |
| Animal (MCD diet: methionine-choline deficient) | Restored normal Bacteroides levels; shifted Erysipelotrichaceae from Allobaculum → Turicibacter; overall altered gut microbiota disrupted by MCD diet | Potential mitigation of diet-induced dysbiosis; supports metabolic homeostasis in liver disease model [39,40] | |
| Animal (Male db/db mice) | ↑ SCFA-producing/anti-inflammatory taxa (Parabacteroides, Oscillibacter, Prevotellaceae); ↓ Anaerotruncus, Lachnospiraceae; correlated with ALT/AST | Anti-inflammatory effects via SCFAs; improved liver markers [41-44] | |
| Human (T2D patients) | ↑ Akkermansia vs metformin; no consistent alpha/beta diversity change in RCTs | Correlate with improved gut-barrier taxa; clinical significance unclear [45] | |
| Semaglutide | Animal (obesity / high-fat diet) | ↑ Akkermansia, Faecalibaculum, Allobaculum; restored dysbiotic taxa; often times reduced diversity | Reduced weight gain, enhanced glucose tolerance, improved gut-barrier integrity [46.47] |
| Animal (PCOS model) | ↑ Helicobacter (negatively correlated with body weight) | Weight loss correlated with microbiome shift [48,49] | |
| Animal (C57BL/6 mice, neurobehavioral model) | Modulates gut microbiota | Reduces hippocampal neuroinflammation; promotes neurogenesis via insulin/GLP-1 pathway; potential antidepressant and anxiolytic effects [50,51] | |
| Dulaglutide | Animal models | ↑ Bacteroides, Akkermansia, Ruminococcus | Improved metabolic patterns (SCFA, gut barrier support) [52,53] |
| Human studies | Limited data; some ↑ Lactobacillus; no significant changes after 1 week, but microbial abundance decreased after 48 weeks in newly diagnosed T2D patients | Metabolic effects observed; microbial effects underpowered; long-term microbiota modulation [54,55] | |
| Exenatide / Exendin-4 | Animal models | ↑ Akkermansia, Barnesiella, Ruminococcus; ↓ dysbiotic taxa | Improved metabolic parameters associated with microbiota shifts [56,57] |
| Human observations | ↑ Coprococcus, Bifidobacterium in small cohorts; otherwise, inconsistent | Limited clinical evidence for direct microbiota mediation [58] | |
| GLP-1Ras (general) | Mixed clinical observations | Some ↑ Akkermansia, Roseburia, Faecalibacterium; SCFA-producing genera enriched; human diversity often unchanged | Suggestive links to glucose tolerance and inflammation regulation; causality not confirmed [59,60] |
| Observational (Human patients) | Increased abundance of Faecalibacterium prausnitzii, negatively correlated with fasting blood glucose levels. | Improved glycemic control; suggests microbiota influences drug responsiveness [61,62] | |
| Observational (Human cohort) | Higher abundance of Roseburia associated with reduced obesity and dyslipidemia; Prevotella/Bacteroides ratio positively associated with obesity, with Prevotella linked to insulin resistance. | Improved metabolism [63,64] | |
| Inflammatory / extra-metabolic models | Animal inflammatory models | Shifts toward more beneficial taxa (SCFA-producing Firmicutes); ↓ pathogenic taxa like Staphylococcus | Enhanced anti-inflammatory activity (IL-22), improved gut barrier; causality in humans unproven to date [65,66] |
Abbreviations: GLP-1: Glucagon-Like Peptide-1; HFD: High-Fat Diet; T2D: Type 2 Diabetes; MCD: Methionine-Choline Deficient; SCFA: Short-Chain Fatty Acids; ALT: Alanine Aminotransferase; AST: Aspartate Aminotransferase; PCOS: Polycystic Ovary Syndrome; IL-22: Interleukin-22.
Moreover, high-resolution taxonomic data as shown in Table 2 [69-93], reveal that liraglutide not only boosts but also maintains the levels of SCFA-producing genera within Bacteroidota (Bacteroides, Alistipes, Parabacteroides, Butyricimonas) and Bacillota (Lactobacillus, Allobaculum, Clostridium, Oscillospira) [36]. At the same time, liraglutide decreases the occurrence of potentially harmful taxa such as Staphylococcus, Anaerotruncus, and Flavonifractor. Importantly, the notable decline in Prevotella_9 is particularly significant given the known link between microbiota dominated by Prevotella and insulin resistance (IR). This highlights the metabolic consequences of the microbial changes induced by GLP-1Ras [67,68].
Table 2: Liraglutide-driven changes in gut microbial taxa.
| Phylum | Genus | Change with Liraglutide | Functional/Metabolic Relevance |
| Bacteroidota | Bacteroides | ↑ Increased / Maintained | SCFA production; glucose homeostasis and gut barrier support [69,70] |
| Alistipes | ↑ Increased / Maintained | Acetate/propionate production; improved insulin sensitivity [71,72] | |
| Parabacteroides | ↑ Increased / Maintained | SCFA production; anti-inflammatory, insulin-sensitizing [73,74] | |
| Butyricimonas | ↑ Increased / Maintained | Butyrate producer; intestinal and metabolic health [75,76] | |
| Bacillota (Firmicutes) | Lactobacillus | ↑ Increased / Maintained | Gut barrier integrity; metabolic signaling (species-dependent) [77-79] |
| Allobaculum | ↑ Increased /Maintained | Linked to leanness and improved insulin sensitivity [80,81] | |
| Clostridium | ↑ Increased / Maintained | Butyrate-producing taxa; gut barrier and anti-inflammatory effects [82,83] | |
| Oscillospira | ↑Increased / Maintained | Associated with leanness and favorable metabolic profile [84,85] | |
| Bacillota | Staphylococcus | ↓ Decreased | Opportunistic genus; dysbiosis and inflammation [86,87] |
| Anaerotruncus | ↓ Decreased | Associated with metabolic dysfunction and inflammation [88,89] | |
| Flavonifractor | ↓ Decreased | Pro-inflammatory; linked to insulin resistance [90,91] | |
| Bacteroidota | Prevotella_9 | ↓ Markedly Decreased | Prevotella dominance associated with insulin resistance [92,93] |
In addition to its function in regulating metabolism, GLP-1 signaling has a notable impact on anti-inflammatory mechanisms and cytoprotection in the gut. When GLP-1 receptors (GLP-1Rs) on intraepithelial lymphocytes (IELs) are activated, there is a decrease in pro-inflammatory cytokine levels, a reduction in the expression of interferon-stimulated genes (ISGs), and a lower rate of apoptosis in epithelial cells [94,95]. In an experiment with mice administered anti-CD3, treatment with exenatide results in a significant drop in levels of interferon gamma (IFNG) and ISG expression in intestinal epithelial cells (IECs), as well as a decrease in crypt cell apoptosis; these outcomes are not observed in mice lacking GLP-1R [96]. These results highlight the crucial importance of GLP-1R signaling in immune cells for sustaining gut homeostasis. In models of colitis, liraglutide successfully diminishes inflammation, maintains crypt structure, and restricts leukocyte infiltration, thus showcasing the protective benefits of GLP-1RAs on intestinal health [97,98].
Furthermore, semaglutide (Table 1) shows distinct microbial profiles, featuring Akkermansia, Faecalibaculum, and Allobaculum, along with improved glucose tolerance and enhanced gut barrier integrity [46,47]. Certain research outcomes are notable, indicating a decrease in microbial diversity, which implies that functional remodeling may play a more significant role than diversity metrics in influencing therapeutic effectiveness. Changes in the microbiome in models of polycystic ovary syndrome (PCOS) have been linked to weight reduction, while studies on neurobehavioral effects have revealed lower inflammation in the hippocampus and increased neurogenesis, thereby supporting the involvement of GLP-1RAs in the GBA [48-51].
More importantly, clinical observations align closely with these preliminary findings. Short-term treatment with liraglutide positively impacts the microbiome, while prolonged dulaglutide treatment leads to gradual changes in the microbial community, mirroring the slow and steady progress in glycemic control, weight loss, and reduction of inflammation [99,100]. It's important to recognize that the baseline composition of the microbiome affects treatment outcomes: a greater presence of Bacteroides dorei and Roseburia inulinivorans is linked to more pronounced decreases in HbA1c, while an increase in Prevotella copri may hinder treatment success [26,101]. Multiple clinical studies consistently show a rise in key strains that produce SCFAs, such as Faecalibacterium prausnitzii and Roseburia, which are inversely related to fasting glucose levels and dyslipidemia [102,103].
Interestingly, research from retrospective studies suggests that GLP-1RAs not only assist but also improve the quality of life for individuals with inflammatory bowel disease (IBD), especially those who have T2D, underscoring the drugs' dual role in metabolic and immune modulation [104,105]. In studies involving mouse models of inflammation, GLP-1RAs facilitate the proliferation of Firmicutes that produce SCFAs and generate Interleukin-22 (IL-22), while also suppressing harmful bacteria like Staphylococcus, which aligns with the observed reduction in epithelial cell death and the activation of the interferon signaling pathway [106,107].
Lastly, GLP-1RAs activate a bidirectional regulatory system that interlinks host metabolism with the immune response in the intestines and the ecology of gut bacteria. The metabolic byproducts produced by bacteria, such as SCFAs, enhance the release of natural GLP-1, while GLP-1RAs alter signaling pathways, acid responses, immune reactions, and the composition of gut microorganisms, leading to increasingly robust therapeutic feedback mechanisms [108,109]. Studies suggest that the gut microbiome acts not merely as a passive entity but as an active biological influencer of GLP-1RA effectiveness, providing compelling support for microbiome-targeted precision therapies for conditions such as T2D and IBD [110,111].
The Gut Microbiome as A Modulator Of GLP-1-Driven Neuroendocrine Signaling
Signals originating from the gut microbiome have a significant effect on neuroendocrine function via several interconnected pathways that correspond with the known mechanisms of GLP-1Ras [112-114]. These pathways include the modulation of gut hormone release, neural communication between the gut and brain, immune system signaling, and metabolic networks in the host. Metabolites that derive from the microbiota, especially SCFAs and secondary bile acids, activate G-protein-coupled receptors such as free fatty acid receptor 2 (FFAR2) and free fatty acid receptor 3 (FFAR3), as well as Takeda G-protein-coupled receptor 5 (TGR5), which are expressed by enteroendocrine L cells [115,116]. Activating these receptors leads to the release of endogenous GLP-1 and other hormones that regulate appetite, including peptide YY (PYY) and serotonin, which in turn impact appetite, glucose regulation, and energy balance [117,118]. Additionally, SCFAs are vital in determining the differentiation, quantity, and secretory capabilities of enteroendocrine cells, which affects both the baseline and stimulus-induced neuroendocrine responses [119,120].
In addition to hormone release, signals from the microbiome influence GBA via vagal afferent pathways that connect to the brainstem and hypothalamic areas responsible for feeding behavior, reward processing, and metabolic control—neural circuits that largely overlap with networks responsive to GLP-1 [121,122]. Evidence supporting this relationship comes from studies on mice treated with antibiotics, which demonstrate that the disruption of gut microbiota leads to increased GLP-1 levels in circulation along with decreased locomotor activity [123,124]. Interestingly, this reduced movement phenotype can be restored by blocking the GLP-1R pharmacologically or by disrupting subdiaphragmatic vagal signaling [123]. On the other hand, directly stimulating GLP-1R in brain regions that send signals through the vagus nerve is enough to trigger reduced locomotor activity, offering practical evidence for a microbiome-GLP-1-vagal pathway in the regulation of neurobehavior [123,125]. Reintroducing specific commensal species, such as Lactobacillus reuteri and Bacteroides thetaiotaomicron, into mice lacking gut microbiota lowers elevated GLP-1 levels and normalizes locomotor activity, thereby establishing a direct connection between microbial composition and GLP-1-related neuroendocrine effects.
Together, these results further support the idea that certain microbial taxa are closely associated with neuroendocrine functions related to GLP-1. For example, specific taxa in the Bacteroidetes phylum, especially Bacteroides species, have been found to affect GLP-1 secretion through the generation of SCFAs like acetate and propionate [126,127]. These metabolites promote the activity of enteroendocrine L-cells via the signaling pathways of free fatty acid receptors 2 and 3 (FFAR2/3), establishing a mechanistic connection between microbial metabolism and the regulation of gut hormones in the host [128,129]. Conversely, some Firmicutes, including butyrate-producing species like Faecalibacterium, Roseburia, and Eubacterium, are known to enhance the function of enteroendocrine cells and maintain barrier integrity, which in turn has an indirect effect on GLP-1 secretion and neuroendocrine activity [130,131]. Additionally, Lactobacilli have been shown to affect bile acid metabolism and vagal signaling, which further influences GLP-1 secretion and associated behavioral patterns [132,133]. The rise in specific microbial species, particularly Akkermansia muciniphila, has been linked to positive metabolic characteristics and increased secretion of gut hormones such as GLP-1, with stronger associations noted in both human and animal research [134,135].
In essence, the overall results indicate that specific microbial communities and functional groups, instead of overall microbial diversity, are crucial in modulating GLP-1-related neuroendocrine activities. This underscores the significance of microbial composition in governing brain-gut interactions through GLP-1-mediated pathways concerning the management of physiological responses. In this regard, Figure 1 shows that certain gut microbes produce metabolites that stimulate L cells to release GLP-1, PYY (Peptide YY), and serotonin, helping regulate appetite, metabolism, and energy balance while sending signals to the brain via the vagus nerve. These effects are shaped by specific microbial taxa rather than overall diversity, highlighting how targeted changes in the microbiome can influence gut-brain communication.
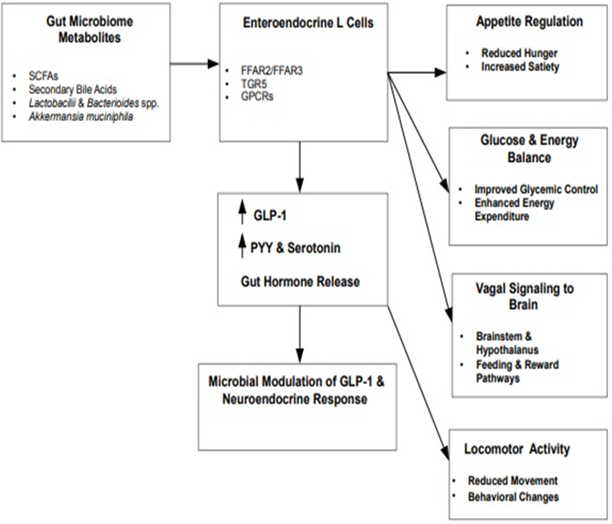
Figure 1: Gut microbiome modulation of GLP-1–driven neuroendocrine signaling. The gut microbiome generates a range of bioactive metabolites, including short-chain fatty acids (SCFAs) and secondary bile acids, which can act on enteroendocrine L cells via specific G-protein-coupled receptors, notably free fatty acid receptors 2 and 3 (FFAR2/3) and Takeda G-protein-coupled receptor 5 (TGR5). Activation of these signaling pathways promotes the secretion of gut-derived hormones such as glucagon-like peptide-1 (GLP-1), peptide YY (PYY), and serotonin. Collectively, these hormones regulate appetite, glucose homeostasis, and whole-body energy balance, while also influencing central neuroendocrine processes. In parallel, microbiome-derived signals engage vagal afferent pathways that relay information from the gut to the brainstem and hypothalamus, thereby integrating peripheral metabolic cues with neural circuits governing feeding behavior, reward processing, and locomotor activity. The figure illustrates how specific microbial metabolites and defined microbial taxa, rather than overall microbial diversity, contribute to brain–gut communication through GLP-1-mediated neuroendocrine signaling pathways.
GLP-1 and Microbiota: Linking the Brain and Adipose Tissue
The GLP-1RAs demonstrate strong impacts on all body metabolic functions. The treatment of GLP-1RA alters the composition of gut microbes. The treatment increases the beneficial microbial groups, which include Roseburia, Faecalibacterium prausnitzii, and Akkermansia [19]. Additionally, the treatment decreases the harmful obesity-related gut bacteria pattern, which shows high Firmicutes-to-Bacteroidetes ratios. The changes in gut bacteria lead to increased production of SCFAs such as acetate, propionate, and butyrate, which act on adipocytes through free fatty acid receptors 41 and 43 (GPR41/43) [19,136]. SCFAs induce white adipose tissue (WAT) to develop beige fat through increased expression of thermogenic and mitochondrial regulators, which include uncoupling protein 1 (UCP1) and peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC-1α) [137-139]. In other words, WAT activation results in higher energy expenditure and better metabolic flexibility.
Microbiota changes induced by GLP-1RA treatment initiate two interlinked effects. These begin with alterations in SCFA signaling and subsequently diverge into bile acid-mediated pathways that modulate farnesoid X receptor (FXR) and Takeda G-protein-coupled receptor 5 (TGR5) activity [19 136]. Through these mechanisms, bile acids regulate hepatic lipid distribution and enhance thermogenic activity in AT. Bile acid-derived signals integrate with microbial metabolites to promote beige adipocyte recruitment, thereby supporting systemic lipid homeostasis [140,141]. Concurrently, intestinal signals are relayed through the GBA, which GLP-1RAs engage via two principal routes: vagal afferent signaling and hypothalamic circuits [142,143]. Together, these pathways coordinate appetite regulation, nutrient sensing, and sympathetic nervous system activity, ultimately shaping energy expenditure and fat storage. The convergence of bile acid-FXR/TGR5 signaling, microbial metabolite-driven adipocyte gene programs, and central neuroendocrine control establishes a microbiota-gut-brain-adipose axis through which GLP-1RAs support healthy AT remodeling, WAT beiging, and overall metabolic balance [144,145]. This framework highlights that GLP-1RAs exert metabolic benefits extending beyond glucose regulation, acting as a mechanistic bridge between gut microbial dynamics and the integrated control of AT function and whole-body energy homeostasis [146,147]. In the context, Figure 2 shows that GLP-1RA-driven microbiota changes coordinate SCFA and bile acid signaling, promoting WAT beiging and gut-brain axis-mediated energy balance.
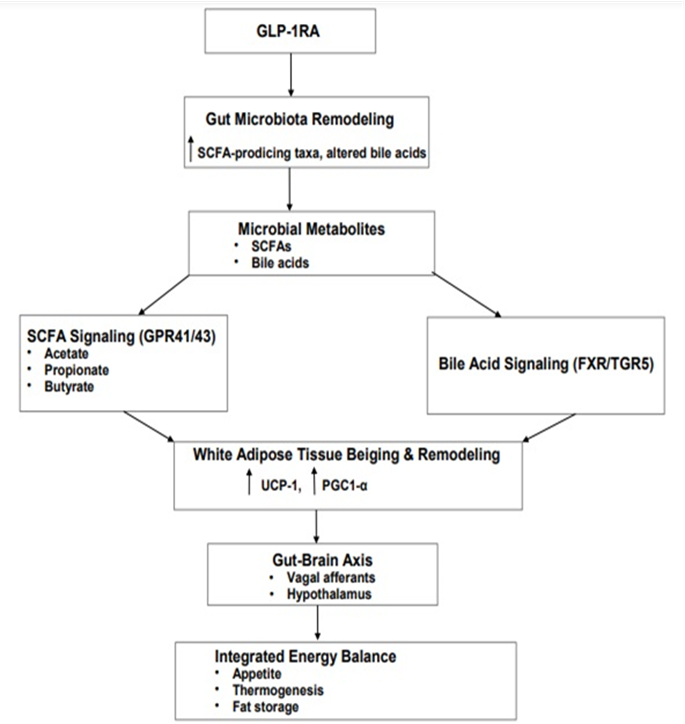
Figure 2: Glucagon-like peptide-1 receptor agonist–mediated microbiota-gut-brain-adipose axis. Glucagon-like peptide-1 receptor agonists (GLP-1RAs) remodel the gut microbiota, increasing short-chain fatty acid (SCFA) production and altering bile acid signaling. SCFAs act on adipocytes via G-protein-coupled receptors 41 and 43 (GPR41/43) to promote white adipose tissue (WAT) beiging and thermogenic gene expression. In parallel, microbiota-modified bile acids activate the farnesoid X receptor (FXR) and Takeda G-protein-coupled receptor 5 (TGR5), regulating hepatic lipid metabolism and enhancing adipose thermogenesis. Intestinal signals are relayed through the gut-brain axis via vagal and hypothalamic pathways, coordinating appetite, energy expenditure, and fat storage. Together, these mechanisms establish an integrated microbiota-gut-brain-adipose axis underlying the metabolic benefits of GLP-1RAs.
GLP-1, Gut Microbiota, and Sarcopenia: Mechanistic Links
GLP-1RAs help preserve skeletal muscle homeostasis by reshaping the intestinal microbiome and its metabolic outputs, offering a novel therapeutic avenue for sarcopenia [148-150]. GLP-1RA treatment alters gut microbial composition, increasing the abundance of beneficial SCFA-producing bacteria such as Roseburia and Faecalibacterium prausnitzii, while reducing dysbiosis-associated taxa [45,47,52,54,59,68]. SCFAs-including acetate, propionate, and butyrate-activate GPR41 and GPR43 receptors expressed on myocytes and engage metabolic signaling pathways such as AMP-activated protein kinase (AMPK) and peroxisome proliferator-activated receptor (PPAR) networks, thereby promoting mitochondrial biogenesis, fatty acid oxidation, and protein synthesis [151-153].
GLP-1RA-induced microbial shifts also attenuate chronic low-grade systemic inflammation by reducing pro-inflammatory cytokines, including tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6), which supports muscle protein integrity and recovery [154,155]. In parallel, microbial bile acid transformation improves insulin sensitivity, enhancing nutrient uptake and metabolic efficiency in myocytes [156,157]. Activation of the gut-brain-muscle axis occurs as GLP-1 signaling engages dual circuits linking vagal afferent inputs to hypothalamic pathways, coordinating neuroendocrine outputs such as insulin-like growth factor-1 (IGF-1) signaling and sympathetic nervous system (SNS) activity that help maintain muscle mass and strength during aging [158-160].
Collectively, the GLP-1-microbiota-metabolite-gut-brain-muscle network integrates metabolic, inflammatory, and neuroendocrine pathways to support skeletal muscle maintenance, enhance functional strength, and reduce sarcopenia risk, highlighting the potential of GLP-1-based and microbiota-targeted strategies in the management of aging-related and metabolic disorders.
Future Directions
The gut-brain-microbiome axis encompasses interconnected metabolic pathways that converge on central systems regulating energy balance through insulin and leptin signaling. Brain IR is now recognized as a key contributor to obesity, T2D, and cognitive dysfunction, while leptin resistance disrupts hypothalamic satiety pathways and promotes maladaptive eating behaviors. Although microbiome modulation improves peripheral insulin sensitivity and reduces inflammatory burden, evidence for restoration of central insulin and leptin sensitivity remains limited. Microbiota-derived metabolites influence neuroinflammation, blood-brain barrier integrity, vagal signaling, and hypothalamic circuitry, suggesting that enhancing endogenous GLP-1 may complement pharmacologic GLP-1R agonism by engaging both peripheral and central metabolic pathways, warranting more evidence-based mechanistic insights in the future.
Conclusion
GLP-1RAs influence metabolic processes through pathways that integrate gut microbiome dynamics with immune responses, neural signaling, and peripheral metabolic tissues. The evidence presented in this review shows that GLP-1RAs consistently remodel gut microbial communities toward SCFA-producing and anti-inflammatory taxa, strengthening endogenous GLP-1 signaling, supporting gut-brain communication, and promoting healthy AT remodeling. Emerging findings further suggest that microbiome-mediated pathways may help preserve skeletal muscle during pharmacologically induced weight loss, raising important considerations regarding sarcopenia. The originality of this review lies in its unified framework, which positions the gut microbiome as an active biological mediator of GLP-1RA action across interconnected systems spanning the gut, brain, AT, and skeletal muscle. By linking microbial activity to lean mass preservation through integrated neuroendocrine and immune mechanisms, this work provides a foundation for personalized GLP-1–based therapeutic strategies that leverage microbiome insights to improve treatment efficacy and long-term metabolic health.
Declarations
Acknowledgments
The author would like to thank Dhrubajyoti Chattopadhyay for his critical review of this work.
Funding
None.
Conflict of Interest
The authors declare no conflict of interest.
References
- He X, Zhao W, Li P, Zhang Y, Li G, et al. (2025). Research progress of GLP-1RAs in the treatment of type 2 diabetes mellitus. Front Pharmacol. 15:1483792.
Publisher | Google Scholor - Shao Q, Xiong J, Wu J, Mao J, Hu Q. (2026). Research progress on oral glucagon-like peptide-1 receptor agonists in the treatment of diabetes mellitus type 2. Front Mol Biosci. 12:1729904.
Publisher | Google Scholor - Kaye AD, Lien N, Vuong C, Schmitt MH, Soorya Y, Abubakar BA, et al. (2024). Glucagon-Like Peptide-1 Receptor Agonist Mediated Weight Loss and Diabetes Mellitus Benefits: A Narrative Review. Cureus. 16(12):e76101.
Publisher | Google Scholor - Westermeier F, Fisman EZ. (2025). Glucagon like peptide-1 (GLP-1) agonists and cardiometabolic protection: historical development and future challenges. Cardiovasc Diabetol. 24(1):44.
Publisher | Google Scholor - Abu-Nejim H, Becker RC. (2025). Current Perspectives on GLP-1 Agonists in Contemporary Clinical Practice from Science and Mechanistic Foundations to Optimal Translation. Curr Atheroscler Rep. 27(1):99.
Publisher | Google Scholor - Beutler LR. (2026). GLP-1 physiology and pharmacology along the gut-brain axis. J Clin Invest. 136(2):e194744.
Publisher | Google Scholor - Zhao X, Wang M, Wen Z, Lu Z, Cui L, et al. (2021). GLP-1 Receptor Agonists: Beyond Their Pancreatic Effects. Front Endocrinol (Lausanne). 12:721135.
Publisher | Google Scholor - Stanimirov B, Đanić M, Pavlović N, Zaklan D, Lazarević S, et al. (2025). Gut-Brain Axis and Bile Acid Signaling: Linking Microbial Metabolism to Brain Function and Metabolic Regulation. Int J Mol Sci. 26(24):12167.
Publisher | Google Scholor - Cryan JF, O'Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, et al. (2019). The Microbiota-Gut-Brain Axis. Physiol Rev. 99(4):1877-2013.
Publisher | Google Scholor - Gruber T, Lechner F, Krieger JP, García-Cáceres C. (2025). Neuroendocrine gut-brain signaling in obesity. Trends Endocrinol Metab. 36(1):42-54.
Publisher | Google Scholor - Reiss AB, Gulkarov S, Lau R, Klek SP, Srivastava A, et al. (2025). Weight Reduction with GLP-1 Agonists and Paths for Discontinuation While Maintaining Weight Loss. Biomolecules. 15(3):408.
Publisher | Google Scholor - Moiz A, Filion KB, Tsoukas MA, Yu OHY, Peters TM, et al. (2025). The expanding role of GLP-1 receptor agonists: a narrative review of current evidence and future directions. EClinicalMedicine. 86:103363.
Publisher | Google Scholor - Kim JA, Yoo HJ. (2025). Exploring the Side Effects of GLP-1 Receptor Agonist: To Ensure Its Optimal Positioning. Diabetes Metab J. 49(4):525-541.
Publisher | Google Scholor - Ayoub M, Chela H, Amin N, Hunter R, Anwar J, et al. (2025). Pancreatitis Risk Associated with GLP-1 Receptor Agonists, Considered as a Single Class, in a Comorbidity-Free Subgroup of Type 2 Diabetes Patients in the United States: A Propensity Score-Matched Analysis. J Clin Med. 14(3):944.
Publisher | Google Scholor - Aimelet V, Holst JJ. (2026). Pharmacological intervention: Challenges and promising outcomes for fat loss and preservation of lean body mass in the treatment of overweight and type 2 diabetes. Diabetes Obes Metab. 28(2):803-816.
Publisher | Google Scholor - Balakrishnan R, Kang SI, Lee JY, Rho YK, Kim BK, et al. (2024). Gut Microbiota-Immune System Interactions in Health and Neurodegenerative Diseases: Insights into Molecular Mechanisms and Therapeutic Applications. Aging Dis. 16(6):3421-3452.
Publisher | Google Scholor - Zhao S, Fu D, Lin Y, Sun X, Wang X, et al. (2025). The role of the microbiome on immune homeostasis of the host nervous system. Front Immunol. 16:1609960.
Publisher | Google Scholor - Almansour N, Al-Rashed F, Choudhry K, Alqaderi H, Sindhu S, et al. (2025). Gut microbiota: a promising new target in immune tolerance. Front Immunol. 16:1607388.
Publisher | Google Scholor - Gofron KK, Wasilewski A, Małgorzewicz S. (2025). Effects of GLP-1 Analogues and Agonists on the Gut Microbiota: A Systematic Review. Nutrients. 17(8):1303.
Publisher | Google Scholor - Münte E, Hartmann P. (2025). The Role of Short-Chain Fatty Acids in Metabolic Dysfunction-Associated Steatotic Liver Disease and Other Metabolic Diseases. Biomolecules. 15(4):469.
Publisher | Google Scholor - Zeng Y, Wu Y, Zhang Q, Xiao X. (2024). Crosstalk between glucagon-like peptide 1 and gut microbiota in metabolic diseases. mBio. 15(1):e0203223.
Publisher | Google Scholor - Kalkan AE, BinMowyna MN, Raposo A, Ahmad MF, Ahmed F, et al. (2025). Beyond the Gut: Unveiling Butyrate's Global Health Impact Through Gut Health and Dysbiosis-Related Conditions: A Narrative Review. Nutrients. 17(8):1305.
Publisher | Google Scholor - Shin Y, Han S, Kwon J, Ju S, Choi TG, et al. (2023). Roles of Short-Chain Fatty Acids in Inflammatory Bowel Disease. Nutrients. 15(20):4466.
Publisher | Google Scholor - Liu SH, Yang XF, Liang L, Song BB, Song XM, et al. (2025). Regulatory mechanisms of the gut microbiota-short chain fatty acids signaling axis in slow transit constipation and progress in multi-target interventions. Front Microbiol. 16:1689597.
Publisher | Google Scholor - Trapanese V, Dagostino A, Natale MR, Giofrè F, Vatalaro C, et al. (2025). Bidirectional Interactions Between the Gut Microbiota and Incretin-Based Therapies. Life (Basel). 15(6):843.
Publisher | Google Scholor - Tsai CY, Lu HC, Chou YH, Liu PY, Chen HY, et al. (2022). Gut Microbial Signatures for Glycemic Responses of GLP-1 Receptor Agonists in Type 2 Diabetic Patients: A Pilot Study. Front Endocrinol (Lausanne). 12:814770.
Publisher | Google Scholor - Kanbay M, Al-Shiab R, Shah E, Ozbek L, Guldan M, et al. (2025). Gut microbiota modulation in GLP-1RA and SGLT-2i therapy: clinical implications and mechanistic insights in type 2 diabetes. Clin Kidney J. 18(12):sfaf351.
Publisher | Google Scholor - Pantazopoulos D, Gouveri E, Papazoglou D, Papanas N. (2025). GLP-1 receptor agonists and sarcopenia: Weight loss at a cost? A brief narrative review. Diabetes Res Clin Pract. 229:112924.
Publisher | Google Scholor - Abildinova GZ, Benberin VV, Vochshenkova TA, Afshar A, Mussin NM, et al. (2024). The gut-brain-metabolic axis: exploring the role of microbiota in insulin resistance and cognitive function. Front Microbiol. 15:1463958.
Publisher | Google Scholor - Bostick JW, Schonhoff AM, Mazmanian SK. (2022). Gut microbiome-mediated regulation of neuroinflammation. Curr Opin Immunol. 76:102177.
Publisher | Google Scholor - Loh JS, Mak WQ, Tan LKS, Ng CX, Chan HH, et al. (2024). Microbiota-gut-brain axis and its therapeutic applications in neurodegenerative diseases. Signal Transduct Target Ther. 9(1):37.
Publisher | Google Scholor - Wang L, Qi G, Shi Y, Ma Y, Gao J. (2025). A systematic longitudinal study of microbiome: integrating temporal-spatial dimensions with causal and deep learning models. BMC Genomics. 26(1):1068.
Publisher | Google Scholor - Gao F, Cheng C, Li R, Chen Z, Tang K, et al. (2025). The role of Akkermansia muciniphila in maintaining health: a bibliometric study. Front Med (Lausanne). 12:1484656.
Publisher | Google Scholor - Bretto E, Urpì-Ferreruela M, Casanova GR, González-Suárez B. (2025). The Role of Gut Microbiota in Gastrointestinal Immune Homeostasis and Inflammation: Implications for Inflammatory Bowel Disease. Biomedicines. 13(8):1807.
Publisher | Google Scholor - Guney-Coskun M, Basaranoglu M. (2024). Interplay of gut microbiota, glucagon-like peptide receptor agonists, and nutrition: New frontiers in metabolic dysfunction-associated steatotic liver disease therapy. World J Gastroenterol. 30(43):4682-4688.
Publisher | Google Scholor - Zhao L, Qiu Y, Zhang P, Wu X, Zhao Z, et al. (2022). Gut microbiota mediates positive effects of liraglutide on dyslipidemia in mice fed a high-fat diet. Front Nutr. 9:1048693.
Publisher | Google Scholor - Shang J, Liu F, Zhang B, Dong K, Lu M, et al. (2021). Liraglutide-induced structural modulation of the gut microbiota in patients with type 2 diabetes mellitus. PeerJ. 9:e11128.
Publisher | Google Scholor - Zhang N, Tao J, Gao L, Bi Y, Li P, et al. (2020). Liraglutide Attenuates Nonalcoholic Fatty Liver Disease by Modulating Gut Microbiota in Rats Administered a High-Fat Diet. Biomed Res Int. 2947549.
Publisher | Google Scholor - Somm E, Montandon SA, Loizides-Mangold U, Gaïa N, Lazarevic V, De Vito C, et al. (2021). The GLP-1R agonist liraglutide limits hepatic lipotoxicity and inflammatory response in mice fed a methionine-choline deficient diet. Transl Res. 227:75-88.
Publisher | Google Scholor - Tsiampali C, Vachliotis ID, Goulas A, Polyzos SA. (2024). Animal studies on glucagon-like peptide-1 receptor agonists and related polyagonists in nonalcoholic fatty liver disease. Hormones (Athens). 23(4):611-619.
Publisher | Google Scholor - Markowiak-Kopeć P, Śliżewska K. (2020). The Effect of Probiotics on the Production of Short-Chain Fatty Acids by Human Intestinal Microbiome. Nutrients. 12(4):1107.
Publisher | Google Scholor - Hizo GH, Rampelotto PH. (2024). The Impact of Probiotic Bifidobacterium on Liver Diseases and the Microbiota. Life (Basel). 14(2):239.
Publisher | Google Scholor - Yan M, Pan D, Chen L, Pang J, Shao Y, et al. (2025). Role of intestinal SCFAs homeostasis in the hepatoprotective effect of Clostridium butyricum in T2DM. NPJ Biofilms Microbiomes. 11(1):206.
Publisher | Google Scholor - Gofron KK, Wasilewski A, Małgorzewicz S. (2025). Effects of GLP-1 Analogues and Agonists on the Gut Microbiota: A Systematic Review. Nutrients. 17(8):1303.
Publisher | Google Scholor - Kant R, Chandra L, Verma V, Nain P, Bello D, et al. (2022). Gut microbiota interactions with anti-diabetic medications and pathogenesis of type 2 diabetes mellitus. World J Methodol. 12(4):246-257.
Publisher | Google Scholor - Duan X, Zhang L, Liao Y, Lin Z, Guo C, et al. (2024). Semaglutide alleviates gut microbiota dysbiosis induced by a high-fat diet. Eur J Pharmacol. 969:176440.
Publisher | Google Scholor - Sun L, Shang B, Lv S, Liu G, Wu Q, et al. (2025). Effects of semaglutide on metabolism and gut microbiota in high-fat diet-induced obese mice. Front Pharmacol. 16:1562896.
Publisher | Google Scholor - Xiong C, Wu J, Ma Y, Li N, Wang X, et al. (2024). Effects of Glucagon-Like Peptide-1 Receptor Agonists on Gut Microbiota in Dehydroepiandrosterone-Induced Polycystic Ovary Syndrome Mice: Compared Evaluation of Liraglutide and Semaglutide Intervention. Diabetes Metab Syndr Obes. 17:865-880.
Publisher | Google Scholor - Carmina E, Longo RA. (2023). Semaglutide Treatment of Excessive Body Weight in Obese PCOS Patients Unresponsive to Lifestyle Programs. J Clin Med. 12(18):5921.
Publisher | Google Scholor - de Paiva IHR, da Silva RS, Mendonça IP, de Souza JRB, Peixoto CA. (2024). Semaglutide Attenuates Anxious and Depressive-Like Behaviors and Reverses the Cognitive Impairment in a Type 2 Diabetes Mellitus Mouse Model Via the Microbiota-Gut-Brain Axis. J Neuroimmune Pharmacol. 19(1):36.
Publisher | Google Scholor - Qi L, Kang H, Zeng F, Zhan M, Huang C, et al. (2025). Gut microbiota mediates semaglutide attenuation of diabetes-associated cognitive decline. Neurotherapeutics. 22(5):e00615.
Publisher | Google Scholor - Liang L, Su X, Guan Y, Wu B, Zhang X, et al. (2024). Correlation between intestinal flora and GLP-1 receptor agonist dulaglutide in type 2 diabetes mellitus treatment-A preliminary longitudinal study. iScience. 27(5):109784.
Publisher | Google Scholor - Mierlan OL, Busila C, Amaritei O, Elena D, Raileanu CR, et al. (2025). Akkermansia muciniphila in Metabolic Disease: Far from Perfect. Int J Mol Sci. 26(23):11602.
Publisher | Google Scholor - Montandon SA, Jornayvaz FR. (2017). Effects of Antidiabetic Drugs on Gut Microbiota Composition. Genes (Basel). 8(10):250.
Publisher | Google Scholor - Fan Y, Liu Y, Shao C, Jiang C, Wu L, et al. (2025). Gut Microbiota-Targeted Therapeutics for Metabolic Disorders: Mechanistic Insights into the Synergy of Probiotic-Fermented Herbal Bioactives. Int J Mol Sci. 26(12):5486.
Publisher | Google Scholor - Khalili L, Park G, Nagpal R, Salazar G. (2024). The Role of Akkermansia muciniphila on Improving Gut and Metabolic Health Modulation: A Meta-Analysis of Preclinical Mouse Model Studies. Microorganisms. 12(8):1627.
Publisher | Google Scholor - Schneeberger M, Everard A, Gómez-Valadés AG, Matamoros S, Ramírez S, et al. (2015). Akkermansia muciniphila inversely correlates with the onset of inflammation, altered adipose tissue metabolism and metabolic disorders during obesity in mice. Sci Rep. 5:16643.
Publisher | Google Scholor - Vergoz V, Jeong D, Hamilton-Williams EE. (2025). Are Bifidobacterium Species Key Players in the Progression of Type 1 Diabetes? A Systematic Review. Endocrinol Diabetes Metab. 8(6):e70120.
Publisher | Google Scholor - Kanbay M, Al-Shiab R, Shah E, Ozbek L, Guldan M, et al. (2025). Gut microbiota modulation in GLP-1RA and SGLT-2i therapy: clinical implications and mechanistic insights in type 2 diabetes. Clin Kidney J. 18(12):sfaf351.
Publisher | Google Scholor - Martinez TM, Wachsmuth HR, Meyer RK, Weninger SN, Lane AI, et al. (2023). Differential effects of plant-based flours on metabolic homeostasis and the gut microbiota in high-fat fed rats. Nutr Metab (Lond). 20(1):44.
Publisher | Google Scholor - Liang L, Rao E, Zhang X, Wu B, Su X, et al. (2023). GLP-1 receptor agonists modulate blood glucose levels in T2DM by affecting Faecalibacterium prausnitzii abundance in the intestine. Medicine (Baltimore). 102(35):e34978.
Publisher | Google Scholor - Kallassy J, Gagnon E, Rosenberg D, Silbart LK, McManus SA. (2023). Strains of Faecalibacterium prausnitzii and its extracts reduce blood glucose levels, percent HbA1c, and improve glucose tolerance without causing hypoglycemic side effects in diabetic and prediabetic mice. BMJ Open Diabetes Res Care. 11(3):e003101.
Publisher | Google Scholor - Singh K, Aulakh SK, Nijjar GS, Singh S, Sandhu APS, et al. (2024). Rebalancing the Gut: Glucagon-Like Peptide-1 Agonists as a Strategy for Obesity and Metabolic Health. Cureus. 16(7):e64738.
Publisher | Google Scholor - Zhang L, Wang P, Huang J, Xing Y, Wong FS, et al. (2024). Gut microbiota and therapy for obesity and type 2 diabetes. Front Endocrinol (Lausanne). 15:1333778.
Publisher | Google Scholor - Nireeksha, Maniangat Luke A, Kumari N S, Hegde MN, Hegde NN. (2025). Metabolic interplay of SCFA's in the gut and oral microbiome: a link to health and disease. Front Oral Health. 6:1646382.
Publisher | Google Scholor - Van K, Burns JL, Monk JM. (2024). Effect of Short-Chain Fatty Acids on Inflammatory and Metabolic Function in an Obese Skeletal Muscle Cell Culture Model. Nutrients. 16(4):500.
Publisher | Google Scholor - Zhang Y, Yang X, Yang P, Sun H, Chen L, et al. (2025). Effect of liraglutide on the dysglycemia, inflammation, and gut microbiota in prediabetic KKay mice. Front Pharmacol. 16:1714859.
Publisher | Google Scholor - Shang J, Liu F, Zhang B, Dong K, Lu M, et al. (2021). Liraglutide-induced structural modulation of the gut microbiota in patients with type 2 diabetes mellitus. PeerJ. 9:e11128.
Publisher | Google Scholor - Zhang Q, Xiao X, Zheng J, Li M, Yu M, et al. (2018). Featured article: Structure moderation of gut microbiota in liraglutide-treated diabetic male rats. Exp Biol Med (Maywood). 243(1):34-44.
Publisher | Google Scholor - Crudele L, Gadaleta RM, Cariello M, Moschetta A. (2023). Gut microbiota in the pathogenesis and therapeutic approaches of diabetes. EBioMedicine. 97:104821.
Publisher | Google Scholor - Zhou JY, Poudel A, Welchko R, Mekala N, Chandramani-Shivalingappa P, et al. (2019). Liraglutide improves insulin sensitivity in high fat diet induced diabetic mice through multiple pathways. Eur J Pharmacol. 861:172594.
Publisher | Google Scholor - Mashayekhi M, Nian H, Mayfield D, Devin JK, Gamboa JL, et al. (2024). Weight Loss-Independent Effect of Liraglutide on Insulin Sensitivity in Individuals with Obesity and Prediabetes. Diabetes. 73(1):38-50.
Publisher | Google Scholor - Liu J, Qiu H, Zhao J, Shao N, Chen C, et al. (2025). Parabacteroides as a promising target for disease intervention: current stage and pending issues. NPJ Biofilms Microbiomes. 11(1):137.
Publisher | Google Scholor - Sun Y, Nie Q, Zhang S, He H, Zuo S, et al. (2023). Parabacteroides distasonis ameliorates insulin resistance via activation of intestinal GPR109a. Nat Commun. 14(1):7740.
Publisher | Google Scholor - Lee H, An J, Kim J, Choi D, Song Y, et al. (2022). A Novel Bacterium, Butyricimonas virosa, Preventing HFD-Induced Diabetes and Metabolic Disorders in Mice via GLP-1 Receptor. Front Microbiol. 13:858192.
Publisher | Google Scholor - Wu J, Wang K, Wang X, Pang Y, Jiang C. (2021). The role of the gut microbiome and its metabolites in metabolic diseases. Protein Cell. 12(5):360-373.
Publisher | Google Scholor - Dempsey E, Corr SC. (2022). Lactobacillus spp. for Gastrointestinal Health: Current and Future Perspectives. Front Immunol. 13:840245.
Publisher | Google Scholor - Zhang Y, Zhu X, Yu X, Novák P, Gui Q, et al. (2023). Enhancing intestinal barrier efficiency: A novel metabolic diseases therapy. Front Nutr. 10:1120168.
Publisher | Google Scholor - Abdalqadir N, Adeli K. (2022). GLP-1 and GLP-2 Orchestrate Intestine Integrity, Gut Microbiota, and Immune System Crosstalk. Microorganisms. 10(10):2061.
Publisher | Google Scholor - Mashayekhi M, Nian H, Mayfield D, Devin JK, Gamboa JL, et al. (2024). Weight Loss-Independent Effect of Liraglutide on Insulin Sensitivity in Individuals with Obesity and Prediabetes. Diabetes. 73(1):38-50.
Publisher | Google Scholor - Ruy CC, Santos TWD, Pereira QC, Ribeiro ML. (2025). Anthocyanins Modulation of Gut Microbiota to Reverse Obesity-Driven Inflammation and Insulin Resistance. Nutrients. 17(23):3727.
Publisher | Google Scholor - Stoeva MK, Garcia-So J, Justice N, Myers J, Tyagi S, et al. (2021). Butyrate-producing human gut symbiont, Clostridium butyricum, and its role in health and disease. Gut Microbes. 13(1):1-28.
Publisher | Google Scholor - Luo Y, Jin Y, Wang H, Wang G, Lin Y, et al. (2024). Effects of Clostridium tyrobutyricum on Lipid Metabolism, Intestinal Barrier Function, and Gut Microbiota in Obese Mice Induced by High-Fat Diet. Nutrients. 16(4):493.
Publisher | Google Scholor - Chen Y, Wang M. (2021). New Insights of Anti-Hyperglycemic Agents and Traditional Chinese Medicine on Gut Microbiota in Type 2 Diabetes. Drug Des Devel Ther. 15:4849-4863.
Publisher | Google Scholor - Craciun CI, Neag MA, Catinean A, Mitre AO, Rusu A, et al. (2022). The Relationships between Gut Microbiota and Diabetes Mellitus, and Treatments for Diabetes Mellitus. Biomedicines. 10(2):308.
Publisher | Google Scholor - Kobayashi T, Glatz M, Horiuchi K, Kawasaki H, Akiyama H, et al. (2015). Dysbiosis and Staphylococcus aureus Colonization Drives Inflammation in Atopic Dermatitis. Immunity. 42(4):756-66.
Publisher | Google Scholor - Zobel EH, Ripa RS, von Scholten BJ, Rotbain Curovic V, Kjaer A, et al. (2021). Effect of liraglutide on expression of inflammatory genes in type 2 diabetes. Sci Rep. 11(1):18522.
Publisher | Google Scholor - Liu Q, Cai BY, Zhu LX, Xin X, Wang X, et al. (2020). Liraglutide modulates gut microbiome and attenuates nonalcoholic fatty liver in db/db mice. Life Sci. 261:118457.
Publisher | Google Scholor - Chen Y, Shan Y, Wang T, Liu Z, Zhao Z, et al. (2025). The Effect of Semaglutide on Gut Microbiota in Chinese Patients with Type 2 Diabetes Poorly Controlled by Metformin. Diabetes Metab Syndr Obes. 18:3865-3881.
Publisher | Google Scholor - Wang K, Lai W, Min T, Wei J, Bai Y, et al. (2024). The Effect of Enteric-Derived Lipopolysaccharides on Obesity. Int J Mol Sci. 25(8):4305.
Publisher | Google Scholor - Martínez-López YE, Esquivel-Hernández DA, Sánchez-Castañeda JP, Neri-Rosario D, Guardado-Mendoza R, et al. (2022). Type 2 diabetes, gut microbiome, and systems biology: A novel perspective for a new era. Gut Microbes. 14(1):2111952.
Publisher | Google Scholor - Gong J, Zhang Q, Hu R, Yang X, Fang C, et al. (2024). Effects of Prevotella copri on insulin, gut microbiota and bile acids. Gut Microbes. 16(1):2340487.
Publisher | Google Scholor - Zhao L, Chen Y, Xia F, Abudukerimu B, Zhang W, et al. (2018). A Glucagon-Like Peptide-1 Receptor Agonist Lowers Weight by Modulating the Structure of Gut Microbiota. Front Endocrinol (Lausanne). 9:233.
Publisher | Google Scholor - Alharbi SH. (2024). Anti-inflammatory role of glucagon-like peptide 1 receptor agonists and its clinical implications. Ther Adv Endocrinol Metab. 15:20420188231222367.
Publisher | Google Scholor - Kumar V. (2025). GLP-1/GLP-1R axis: from metabolism (obesity and T2DM) to immunity. Open Biol. 15(7):240303.
Publisher | Google Scholor - Wong CK, Yusta B, Koehler JA, Baggio LL, McLean BA, et al. (2022). Divergent roles for the gut intraepithelial lymphocyte GLP-1R in control of metabolism, microbiota, and T cell-induced inflammation. Cell Metab. 34(10):1514-1531.e7.
Publisher | Google Scholor - Saadoun AA, Abdelsattar AH, Elsaid AH, Abdelaleam EA, Abdelkader HK, et al. (2025). Repurposing liraglutide to the management of DSS-induced colitis: a potential for promoting autophagy. Naunyn Schmiedebergs Arch Pharmacol. 398(12):17173-17185.
Publisher | Google Scholor - Wang W, Zhang C, Zhang H, Li L, Fan T, et al. (2023). The alleviating effect and mechanism of GLP-1 on ulcerative colitis. Aging (Albany NY). 15(16):8044-8060.
Publisher | Google Scholor - Bobu AM, Turliuc S, Cucu AI, Onofriescu A, Dascalu CG, et al. (2025). Dulaglutide 1.5 mg Significantly Improves Glycemic Control and Lowers LDL-Cholesterol and Body Weight in Romanian Patients with Type 2 Diabetes. J Clin Med. 14(10):3536.
Publisher | Google Scholor - Kim HS, Kim MJ, Kim HS, Cho YK, Jung CH, et al. (2025). Real-World Evidence of Long-Term Dulaglutide Use: Sustained Glycemic and Weight Improvements Beyond Three Years. Clin Endocrinol (Oxf).
Publisher | Google Scholor - Florêncio GP, Xavier AR, Natal ACC, Sadoyama LP, Pena GDG, et al. (2025). Synergistic Effects of Probiotics and Lifestyle Interventions on Intestinal Microbiota Composition and Clinical Outcomes in Obese Adults. Metabolites. 15(2):70.
Publisher | Google Scholor - Tsai CY, Lu HC, Chou YH, Liu PY, Chen HY, et al. (2022). Gut Microbial Signatures for Glycemic Responses of GLP-1 Receptor Agonists in Type 2 Diabetic Patients: A Pilot Study. Front Endocrinol (Lausanne). 12:814770.
Publisher | Google Scholor - Du Y, He C, An Y, Huang Y, Zhang H, Fu W, et al. (2024). The Role of Short Chain Fatty Acids in Inflammation and Body Health. Int J Mol Sci. 25(13):7379.
Publisher | Google Scholor - Colwill M, Povlsen S, Pollok R, Patel K, Goodhand J, et al. (2025). Glucagon-like peptide-1 (GLP-1) receptor agonists in inflammatory bowel disease: mechanisms, clinical implications, and therapeutic potential. J Crohns Colitis. 19(9):jjaf167.
Publisher | Google Scholor - Migliorisi G, Gabbiadini R, Dal Buono A, Ferraris M, Privitera G, et al. (2025). GLP-1 receptor agonists in IBD: exploring the crossroads of metabolism and inflammation. Front Immunol. 16:1610368.
Publisher | Google Scholor - Sun H, Shu J, Tang J, Li Y, Qiu J, et al. (2024). GLP-1 receptor agonists alleviate colonic inflammation by modulating intestinal microbiota and the function of group 3 innate lymphoid cells. Immunology. 172(3):451-468.
Publisher | Google Scholor - Thin L, Teh WL. (2025). GLP-1R Agonists and Their Therapeutic Potential in Inflammatory Bowel Disease and Other Immune-Mediated Inflammatory Diseases, a Systematic Review of the Literature. Biomedicines. 13(5):1128.
Publisher | Google Scholor - Ganamurali N, Sabarathinam S. (2026). Bidirectional interplay between the gut microbiota and GLP-1 receptor agonists: towards Microbiome-Mediated therapeutics in type 2 diabetes mellitus. J Diabetes Metab Disord. 25(1):44.
Publisher | Google Scholor - Guney-Coskun M, Basaranoglu M. (2024). Interplay of gut microbiota, glucagon-like peptide receptor agonists, and nutrition: New frontiers in metabolic dysfunction-associated steatotic liver disease therapy. World J Gastroenterol. 30(43):4682-4688.
Publisher | Google Scholor - Wang J, Qu J, Ye M, Feng R, Hui X, et al. (2026). Beyond conventional therapies: Gut microbiota modulation and macromolecular drugs in the battle against cardiometabolic diseases. J Pharm Anal. 16(1):101416.
Publisher | Google Scholor - Zeng Y, Wu Y, Zhang Q, Xiao X. (2024). Crosstalk between glucagon-like peptide 1 and gut microbiota in metabolic diseases. mBio. 15(1):e0203223.
Publisher | Google Scholor - Guo TL, Chen Y, Xu HS, McDonough CM, Huang G. (2020). Gut microbiome in neuroendocrine and neuroimmune interactions: The case of genistein. Toxicol Appl Pharmacol. 402:115130.
Publisher | Google Scholor - Wang SZ, Yu YJ, Adeli K. (2020). Role of Gut Microbiota in Neuroendocrine Regulation of Carbohydrate and Lipid Metabolism via the Microbiota-Gut-Brain-Liver Axis. Microorganisms. 8(4):527.
Publisher | Google Scholor - Riehl L, Fürst J, Kress M, Rykalo N. (2024). The importance of the gut microbiome and its signals for a healthy nervous system and the multifaceted mechanisms of neuropsychiatric disorders. Front Neurosci. 17:1302957.
Publisher | Google Scholor - Oteng AB, Liu L. (2023). GPCR-mediated effects of fatty acids and bile acids on glucose homeostasis. Front Endocrinol (Lausanne). 14:1206063.
Publisher | Google Scholor - Masse KE, Lu VB. (2023). Short-chain fatty acids, secondary bile acids and indoles: gut microbial metabolites with effects on enteroendocrine cell function and their potential as therapies for metabolic disease. Front Endocrinol (Lausanne). 14:1169624.
Publisher | Google Scholor - Gou X, Qin L, Wu D, Xie J, Lu Y, et al. (2023). Research Progress of Takeda G Protein-Coupled Receptor 5 in Metabolic Syndrome. Molecules. 28(15):5870.
Publisher | Google Scholor - Movahednasab M, Dianat-Moghadam H, Khodadad S, Nedaeinia R, Safabakhsh S, et al. (2025). GLP-1-based therapies for type 2 diabetes: from single, dual and triple agonists to endogenous GLP-1 production and L-cell differentiation. Diabetol Metab Syndr. 17(1):60.
Publisher | Google Scholor - Pang S, Ren Z, Ding H, Chan P. (2026). Short-chain fatty acids mediate enteric and central nervous system homeostasis in Parkinson's disease: Innovative therapies and their translation. Neural Regen Res. 21(3):938-956.
Publisher | Google Scholor - Silva YP, Bernardi A, Frozza RL. (2020). The Role of Short-Chain Fatty Acids from Gut Microbiota in Gut-Brain Communication. Front Endocrinol (Lausanne). 11:25.
Publisher | Google Scholor - Longo S, Rizza S, Federici M. (2023). Microbiota-gut-brain axis: relationships among the vagus nerve, gut microbiota, obesity, and diabetes. Acta Diabetol. 60(8):1007-1017.
Publisher | Google Scholor - Yu KB, Hsiao EY. (2021). Roles for the gut microbiota in regulating neuronal feeding circuits. J Clin Invest. 131(10):e143772.
Publisher | Google Scholor - Lai TT, Tsai YH, Liou CW, Fan CH, Hou YT, et al. (2024). The gut microbiota modulate locomotion via vagus-dependent glucagon-like peptide-1 signaling. NPJ Biofilms Microbiomes. 10(1):2.
Publisher | Google Scholor - Kellenberger A, Dewal RS, de Wouters d'Oplinter A, Sichert A, Heine M, et al. (2025). Antibiotic-induced gut microbiota depletion enhances glucose tolerance linked to GLP-1 signaling. Front Endocrinol (Lausanne). 16:1684155.
Publisher | Google Scholor - Brierley DI, de Lartigue G. (2022). Reappraising the role of the vagus nerve in GLP-1-mediated regulation of eating. Br J Pharmacol. 179(4):584-599.
Publisher | Google Scholor - Fusco W, Lorenzo MB, Cintoni M, Porcari S, Rinninella E, et al. (2023). Short-Chain Fatty-Acid-Producing Bacteria: Key Components of the Human Gut Microbiota. Nutrients. 15(9):2211.
Publisher | Google Scholor - Zhong Y, Lei Y, Jiang S, Chen D, Wang X, et al. (2025). Advances in understanding the role of gut microbiota in fat deposition and lipid metabolism. J Anim Sci Biotechnol. 16(1):152.
Publisher | Google Scholor - Mishra SP, Karunakar P, Taraphder S, Yadav H. (2020). Free Fatty Acid Receptors 2 and 3 as Microbial Metabolite Sensors to Shape Host Health: Pharmacophysiological View. Biomedicines. 8(6):154.
Publisher | Google Scholor - He J, Zhang P, Shen L, Niu L, Tan Y, et al. (2020). Short-Chain Fatty Acids and Their Association with Signalling Pathways in Inflammation, Glucose and Lipid Metabolism. Int J Mol Sci. 21(17):6356.
Publisher | Google Scholor - Nie K, Ma K, Luo W, Shen Z, Yang Z, et al. (2021). Roseburia intestinalis: A Beneficial Gut Organism from the Discoveries in Genus and Species. Front Cell Infect Microbiol. 11:757718.
Publisher | Google Scholor - Berding K, Vlckova K, Marx W, Schellekens H, Stanton C, et al. (2021). Diet and the Microbiota-Gut-Brain Axis: Sowing the Seeds of Good Mental Health. Adv Nutr. 12(4):1239-1285.
Publisher | Google Scholor - Prete R, Long SL, Gallardo AL, Gahan CG, Corsetti A, et al. (2020). Beneficial bile acid metabolism from Lactobacillus plantarum of food origin. Sci Rep. 10(1):1165.
Publisher | Google Scholor - Baldi S, Turrini M, Cei F, Menicatti M, Bartolucci G, et al. (2026). Bile Acids as Key Mediators of the Gut Microbiota-Immune Axis: Potential Biomarker and Therapeutic Perspectives. Biofactors. 52(1):e70078.
Publisher | Google Scholor - Pellegrino A, Coppola G, Santopaolo F, Gasbarrini A, Ponziani FR. (2023). Role of Akkermansia in Human Diseases: From Causation to Therapeutic Properties. Nutrients. 15(8):1815.
Publisher | Google Scholor - Segers A, de Vos WM. (2023). Mode of action of Akkermansia muciniphila in the intestinal dialogue: role of extracellular proteins, metabolites and cell envelope components. Microbiome Res Rep. 2(1):6.
Publisher | Google Scholor - Singh K, Aulakh SK, Nijjar GS, Singh S, Sandhu APS, et al. (2024). Rebalancing the Gut: Glucagon-Like Peptide-1 Agonists as a Strategy for Obesity and Metabolic Health. Cureus. 16(7):e64738.
Publisher | Google Scholor - Ghesmati Z, Rashid M, Fayezi S, Gieseler F, Alizadeh E, et al. (2023). An update on the secretory functions of brown, white, and beige adipose tissue: Towards therapeutic applications. Rev Endocr Metab Disord. 25(2):279-308.
Publisher | Google Scholor - Alzubi A, Glowacki HX, Burns JL, Van K, Martin JLA, et al. (2025). Dose-Dependent Effects of Short-Chain Fatty Acids on 3T3-L1 Adipocyte Adipokine Secretion and Metabolic Function. Nutrients. 17(3):571.
Publisher | Google Scholor - Li G, Xie C, Lu S, Nichols RG, Tian Y, et al. (2017). Intermittent Fasting Promotes White Adipose Browning and Decreases Obesity by Shaping the Gut Microbiota. Cell Metab. 26(4):672-685.e4.
Publisher | Google Scholor - Liu M, He C, Lv X, Zheng W, Yin J, et al. (2026). Bile acid signaling mediates gut microbiota regulation of host lipid metabolism. Front Microbiol. 16:1696213.
Publisher | Google Scholor - He Y, Shaoyong W, Chen Y, Li M, Gan Y, et al. (2026). The functions of gut microbiota-mediated bile acid metabolism in intestinal immunity. J Adv Res. 80:351-370.
Publisher | Google Scholor - Liu QK. (2024). Mechanisms of action and therapeutic applications of GLP-1 and dual GIP/GLP-1 receptor agonists. Front Endocrinol (Lausanne). 15:1431292.
Publisher | Google Scholor - Liu Z, Yu S, Jin X, Sheng L, YanMu MR, et al. (2025). The Clinical Application of GLP-1RAs and GLP-1/GIP Dual Receptor Agonists Based on Pharmacological Mechanisms: A Review. Drug Des Devel Ther. 19:10383-10409.
Publisher | Google Scholor - Han B, Lv X, Liu G, Li S, Fan J, et al. (2023). Gut microbiota-related bile acid metabolism-FXR/TGR5 axis impacts the response to anti-α4β7-integrin therapy in humanized mice with colitis. Gut Microbes. 15(1):2232143.
Publisher | Google Scholor - Stanimirov B, Đanić M, Pavlović N, Zaklan D, Lazarević S, et al. (2025). Gut-Brain Axis and Bile Acid Signaling: Linking Microbial Metabolism to Brain Function and Metabolic Regulation. Int J Mol Sci. 26(24):12167.
Publisher | Google Scholor - Salehi M, Purnell JQ. (2019). The Role of Glucagon-Like Peptide-1 in Energy Homeostasis. Metab Syndr Relat Disord. 17(4):183-191.
Publisher | Google Scholor - Zheng Z, Zong Y, Ma Y, Tian Y, Pang Y, et al. (2024). Glucagon-like peptide-1 receptor: mechanisms and advances in therapy. Signal Transduct Target Ther. 9(1):234.
Publisher | Google Scholor - González-Luis A, Llinares-Arvelo V, Martínez-Alberto CE, Hernández-Carballo C, Mora-Fernández C, et al. (2025). Glucagon-like peptide-1 receptor agonists and muscle health: potential role in sarcopenia prevention and treatment. Eur J Endocrinol. 193(5):R31-R44.
Publisher | Google Scholor - Gatto A, Liu K, Milan N, Wong S. (2025). The Effects of GLP-1 Agonists on Musculoskeletal Health and Orthopedic Care. Curr Rev Musculoskelet Med. 18(10):469-480.
Publisher | Google Scholor - Pantazopoulos D, Gouveri E, Papazoglou D, Papanas N. (2025). GLP-1 receptor agonists and sarcopenia: Weight loss at a cost? A brief narrative review. Diabetes Res Clin Pract. 229:112924.
Publisher | Google Scholor - Kimura I, Inoue D, Hirano K, Tsujimoto G. (2014). The SCFA Receptor GPR43 and Energy Metabolism. Front Endocrinol (Lausanne). 5:85.
Publisher | Google Scholor - Otten BMJ, Sthijns MMJPE, Troost FJ. (2023). A Combination of Acetate, Propionate, and Butyrate Increases Glucose Uptake in C2C12 Myotubes. Nutrients. 15(4):946.
Publisher | Google Scholor - Réka F, Zsófia B, Ádám B, Péter F. (2025). The Gut-Muscle-Immune Axis in Motion: Mechanistic Synergies of SCFA Metabolism, Exercise, and Microbial Cross-Feeding. Nutrients. 17(23):3786.
Publisher | Google Scholor - Mehdi SF, Pusapati S, Anwar MS, Lohana D, Kumar P, et al. (2023). Glucagon-like peptide-1: a multi-faceted anti-inflammatory agent. Front Immunol. 14:1148209.
Publisher | Google Scholor - Alharbi SH. (2024). Anti-inflammatory role of glucagon-like peptide 1 receptor agonists and its clinical implications. Ther Adv Endocrinol Metab. 15:20420188231222367.
Publisher | Google Scholor - Chen L, Zhang Z, Deng W, Jiang G, Xie D, et al. (2025). Bile acids enhance fat metabolism and skeletal muscle development in Zhijiang duck by modulating gut microbiota. Poult Sci. 104(8):105319.
Publisher | Google Scholor - Al Qassab M, Chaarani N, Hamou A, Harb R, Jradi A, et al. (2026). The Gut Microbiota-Insulin Resistance Axis: Mechanisms, Clinical Implications, and Therapeutic Potential. FASEB Bioadv. 8(1):e70080.
Publisher | Google Scholor - Diz-Chaves Y, Herrera-Pérez S, González-Matías LC, Lamas JA, Mallo F. (2020). Glucagon-Like Peptide-1 (GLP-1) in the Integration of Neural and Endocrine Responses to Stress. Nutrients. 12(11):3304.
Publisher | Google Scholor - Yin Y, Guo Q, Zhou X, Duan Y, Yang Y, et al. (2022). Role of brain-gut-muscle axis in human health and energy homeostasis. Front Nutr. 9:947033.
Publisher | Google Scholor - Goldstein N, McKnight AD, Carty JRE, Arnold M, Betley JN, et al. (2021). Hypothalamic detection of macronutrients via multiple gut-brain pathways. Cell Metab. 33(3):676-687.e5.
Publisher | Google Scholor























