

Review Article
Adipose Tissue Remodeling: The Expanding Biology of GLP-1 Receptor Agonists
H. P. Ghosh Research Center, New Town, Kolkata, West Bengal 700161, India.
*Corresponding Author: Swarup K. Chakrabarti, H. P. Ghosh Research Center, New Town, Kolkata, West Bengal 700161, India.
Citation: Chakrabarti SK. (2026). Adipose Tissue Remodeling: The Expanding Biology of GLP-1 Receptor Agonists, Journal of Clinical Research and Clinical Trials, BioRes Scientia Publishers. 5(1):1-21. DOI: 10.59657/2837-7184.brs.26.067
Copyright: © 2026 Swarup K. Chakrabarti, this is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Received: March 11, 2026 | Accepted: April 03, 2026 | Published: April 13, 2026
Abstract
The rising prevalence of type 2 diabetes (T2D) and cardiovascular diseases (CVDs) emphasizes the need to rapidly develop effective metabolic therapies. Glucagon-like peptide-1 receptor agonists (GLP-1RAs) have gained attention as potent treatments, traditionally known for their ability to suppress appetite and lower blood glucose levels. However, the mechanisms by which GLP-1RAs remodel adipose tissue (AT), promote the conversion of white fat to brown fat, and modulate adipokine secretion remain incompletely understood, representing a key area for further research. Evidence from preclinical studies and early clinical trials suggests that GLP-1RAs can reduce visceral fat, preserve subcutaneous fat, enhance insulin sensitivity, and decrease inflammation, often without substantial weight loss. These effects appear to result from a combination of central and peripheral processes, including immune remodeling of white adipose tissue (WAT), sympathetic activation of brown and beige fat, regulation of thermogenic gene expression, and broader systemic metabolic improvements. This review synthesizes current insights into GLP-1RA–induced AT remodeling, linking immune, neural, transcriptional, and systemic mechanisms. It emphasizes the emerging view of GLP-1RAs as AT-centric immunometabolic regulators, discusses their clinical relevance, and outlines future research directions to enhance metabolic health beyond traditional weight-focused strategies.
Keywords: GLP-1 agonists; adipose remodeling; browning; visceral fat; insulin sensitivity; metabolic health
Introduction
The increasing burden of type 2 diabetes (T2D) and cardiovascular diseases (CVDs) is directly linked to the metabolic effects of obesity, creating an urgent need for effective interventions. GLP-1-directed therapies have emerged among the most powerful approaches to weight loss, primarily through their well-recognized anorexigenic and glucose-lowering effects [1-3]. Adipose tissue (AT) primarily consists of white adipocytes (WA) and serves as the body's main energy reservoir; it retains fatty acids during energy surplus and liberates them in times of energy shortage, while also playing an essential role in the endocrine control of glucose and lipid metabolism [4,5]. Conversely, brown adipose tissue (BAT) releases energy via thermogenesis, and specific WAT deposits can undergo "beiging" influenced by environmental signals or precursor cell maturation [6]. This plasticity makes beige and brown fat highly attractive targets for metabolic interventions, and GLP-1 receptor agonists (GLP-1 RAs) seem to have the potential to impact these depots selectively [7,8]. Despite recent progress, the mechanisms by which GLP-1-based therapies modulate AT architecture, functionality, and secretory profile remain incompletely understood. Until now, research has placed much emphasis on appetite regulation, glycemic control, or overall weight loss, leaving behind a gap in understanding the effects of GLP-1RAs on AT remodeling, WAT browning, BAT activation, and secretion of adipokines-processes that might critically influence systemic metabolism. Preclinical and preliminary clinical data suggest that, beyond simple adipose mass reduction, GLP-1RAs modulate the secretory and inflammatory features of this tissue [9,10]. By enhancing anti-inflammatory, metabolically advantageous adipokines while concurrently reducing pro-inflammatory mediators, GLP-1RAs improve insulin sensitivity, constrain ectopic lipid accumulation, and optimize energy utilization [11-13]. In animal models, GLP-1RAs induce WAT browning and BAT activation, increasing thermogenesis, mitochondrial biogenesis, and oxidative metabolism [14,15]. Early human studies confirm these findings, as reductions in visceral and subcutaneous adipose tissue (VAT and SAT) are accompanied by improvements in AT function [16,17]. Collectively, these observations indicate that GLP-1-based therapies effect both structural and functional changes in AT, promoting a metabolically active and hormonally beneficial phenotype. By placing the physiological significance of beige and brown adipose depots into context and positioning GLP-1RAs as modulators of AT function rather than simply as weight-loss drugs, this review sets a conceptual framework for targeting AT to enhance long-term metabolic health and decrease obesity-associated complications.
Central-Peripheral GLP-1 Signaling as The Nexus of Adipose Tissue Remodeling
Existing studies support the notion that GLP-1RAs influence AT predominantly through multiple indirect pathways that coordinate activities between the central nervous system (CNS), pancreas, liver, skeletal muscle, and immune system, rather than exerting a primary direct effect on adipocytes [18-20]. Table 1. summarizes GLP-1 receptor (GLP-1R) expression and actions across metabolic tissues [21-31]. The high density of GLP-1Rs in the CNS and pancreatic islets represents the two principal mechanisms underlying their action: suppression of appetite via CNS pathways and reduction of autonomic (sympathetic) activity, which together decrease energy intake, while a balanced insulin/ glucagon response mitigates insulin-stimulated lipogenesis [32,33]. These factors collectively reduce lipid entry into AT, resulting in smaller adipocyte size and improved metabolic flexibility. Human imaging and AT biopsy studies further support these observations, showing decreased adipocyte size, improved AT architecture, and increased circulating anti-inflammatory adiponectin, indicating restoration of adipocyte endocrine function upon GLP-1RA treatment [34-36].
Table 1: GLP-1 Receptor Expression and Actions Across Metabolic Tissues.
| Tissue | GLP-1R Level | GLP-1R-Expressing Cells | Key Functions | GLP-1 RA Action | References |
| Brain (CNS) | High, region-specific | Hypothalamic neurons (POMC; AgRP subsets); brainstem neurons (NTS, area postrema) | Appetite control; satiety; energy balance; autonomic regulation | Direct | [21] |
| Pancreatic Islets | High | β-cells (robust); δ-cells; minimal in α-cells | Insulin secretion; glucagon suppression | Direct | [22] |
| Gastrointestinal Tract | Moderate | Enteric neurons; vagal afferents; smooth muscle cells | Gastric emptying; gut–brain signaling | Direct | [23] |
| Adipose Tissue | Low to very low | Adipocytes (low); stromal cells (variable); macrophages (low) | Tissue remodeling; adipokine secretion; inflammation control | Primarily indirect | [24] |
| Liver | Absent to minimal | Non-parenchymal cells? | Hepatic insulin sensitivity; lipid metabolism | Indirect | [25] |
| Skeletal Muscle | Absent | No convincing myocyte expression | Peripheral glucose uptake | Indirect | [26] |
| Heart | Low to moderate | Cardiomyocytes; endothelial cells | Cardioprotection; metabolic support | Direct and indirect | [27] |
| Kidney | Moderate | Proximal tubules; renal vasculature | Natriuresis; renal protection | Direct and indirect | [28] |
| Lung | Low | Airway smooth muscle; endothelium | Airway tone; inflammation modulation | Direct (limited evidence | [29] |
| Immune Cells | Very low, variable | Macrophages; select T cells | Immunomodulation; inflammation resolution | Primarily indirect | [30] |
| Hypothalamic Microglia | Questionable | Functional effects via neuronal signaling | Neuroinflammation regulation; leptin sensitivity | Indirect | [31] |
Abbreviations: GLP-1, glucagon-like peptide-1; GLP-1 RA, glucagon-like peptide-1 receptor agonist; CNS, central nervous system; POMC neurons, pro-opiomelanocortin neurons; AgRP neurons, agouti-related peptide neurons; NTS, nucleus tractus solitarius.
Moreover, at the tissue level, these improvements in peripheral metabolism facilitate AT remodeling. Enhanced hepatic insulin sensitivity reduces de novo lipogenesis and limits the export of triglyceride-rich lipoproteins (primarily very-low-density lipoproteins, VLDL) to peripheral tissues, thereby decreasing substrate availability to AT [37,38]. Simultaneously, increased glucose uptake by skeletal muscle shifts nutrient utilization toward oxidation rather than storage [39]. In AT, reduced macrophage infiltration and decreased inflammatory signaling relieve cytokine-driven suppression of mitochondrial and adipokine gene expression, promoting healthier lipid turnover [40,41]. Clinical studies consistently show a preferential reduction in visceral adipose tissue (VAT), while subcutaneous adipose tissue (SAT) is either preserved or normalized, indicating a shift toward a more metabolically favorable AT distribution rather than indiscriminate AT loss [42,43].
Lastly, depot-specific findings provide further insight. VAT, which is highly sensitive to inflammation and insulin resistance (IR), shows significant reductions, thereby lowering cardiometabolic risk [44,45]. SAT undergoes functional normalization with mild increases in oxidative function, although robust evidence for significant browning is not yet apparent [46,47]. Regarding BAT, direct evidence remains limited to a single study; however, the absence of inflammation and enhanced neuroendocrine activation may confer indirect benefits. Overall, the literature suggests that GLP-1RAs primarily act by modifying the systemic metabolic milieu to induce depot-specific changes in AT distribution and function [48,49]. Figure 1 shows coordinated central and peripheral actions of GLP-1RAs that drive AT remodeling and improve metabolic health.
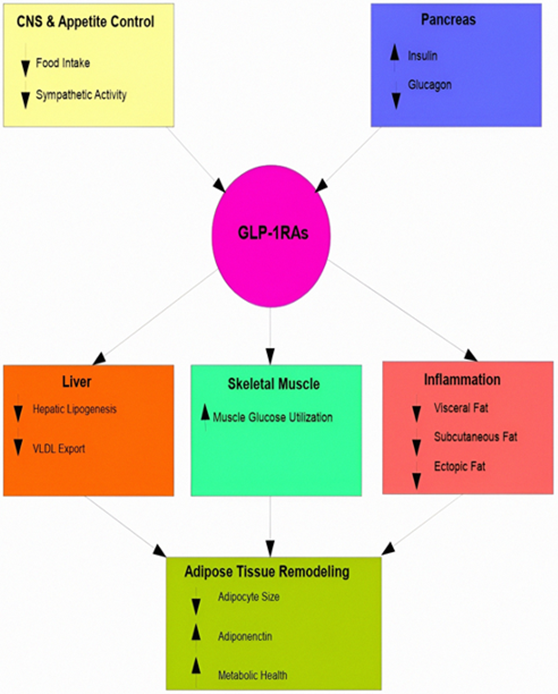
Figure 1: Multisystem actions of GLP-1 receptor agonists in adipose tissue remodeling. GLP-1 receptor agonists (GLP-1RAs) activate synchronized central and peripheral pathways that together transform the biology of adipose tissue. Signaling within the central nervous system (yellow) diminishes food consumption and lowers sympathetic activity, whereas effects on the pancreas (blue) promote insulin secretion and inhibit glucagon release. Actions in the periphery include decreased hepatic lipogenesis and lowered export of very-low-density lipoprotein (VLDL) from the liver (orange), improved glucose usage in skeletal muscle (green), and reduced inflammation along with decreases in visceral, subcutaneous, and ectopic fat stores (red). These coordinated effects lead to remodeling of adipose tissue (lime green), characterized by smaller fat cells, increased levels of adiponectin, and better metabolic health. Upward arrows denote increases, while downward arrows indicate decreases. The color-coded boxes represent the primary organ systems and functional areas affected by GLP-1RAs.
Mechanisms Driving GLP-1 Receptor Agonist-Mediated White-to-Brown Adipose Tissue Conversion
GLP-1RAs, including liraglutide and semaglutide, are known to exert potent anti-inflammatory effects across multiple metabolic tissues, including WAT, the liver, and the vasculature [50,51]. These agents reduce the expression of pro-inflammatory cytokines such as tumor necrosis factor-α (TNF-α) and interleukin-6 (IL-6) and attenuate macrophage accumulation within WAT [52,53]. Chronic low-grade inflammation in WAT is a hallmark of metabolic dysfunction and represents a major barrier to AT browning and thermogenic activation [54,55]. Accordingly, by alleviating this inflammatory state, GLP-1RAs may create a permissive environment that supports adipocyte plasticity [56,57]. The conversion of white adipocytes into beige or brown-like adipocytes is characterized by the activation of key thermogenic gene programs, including uncoupling protein 1 (UCP1), PR domain-containing protein 16 (PRDM16), and peroxisome proliferator-activated receptor γ coactivator-1α (PGC-1α) [58,59]. This thermogenic remodeling is strongly suppressed under conditions of chronic inflammation, oxidative stress, and IR [60,61]. In contrast, anti-inflammatory signaling has been shown to facilitate thermogenic gene expression and promote mitochondrial biogenesis within WAT [62,63]. In this context, the anti-inflammatory actions of GLP-1RAs may indirectly support WAT browning by reducing inhibitory cues that constrain thermogenic pathway activation [64,65]. However, these effects likely represent only one component of a multifactorial regulatory network that also involves neural, endocrine, and metabolic signaling pathways governing the development of brown and beige adipocytes from WAT [66,67].
The anti-inflammatory effects of GLP-1RAs provide a key framework for understanding their influence on AT plasticity; however, inflammation alone is unlikely to explain the full complexity of WAT browning. Thermogenic remodeling of WAT requires coordinated regulation across multiple biological layers, including immune control of the adipose microenvironment, CNS-driven sympathetic signaling, adipocyte-intrinsic metabolic reprogramming, and systemic endocrine cues [68,69]. Emerging evidence suggests that GLP-1 RAs engage each of these regulatory pathways, albeit to varying extents, thereby relieving inhibitory constraints and enabling activation of thermogenic pathways [70,71]. The following sections examine these mechanisms in detail, starting with immunometabolic remodeling of WAT, followed by neural, cellular, mitochondrial, and endocrine pathways that collectively contribute to GLP-1RA-associated adipose browning.
Immunometabolic Remodeling of White Adipose Tissue (WAT)
WAT is a metabolically active tissue whose ability to maintain systemic homeostasis is strongly influenced by immune factors [72,73]. Its cellular composition and inflammatory status are critical determinants of functional capacity. In obesity and metabolic disorders, WAT shows a chronic low-grade inflammatory state with elevated pro-inflammatory cytokines like TNF-α and IL-6 and an increased abundance of M1-like macrophages [74,75]. This inflammatory environment activates stress-responsive signaling pathways, leading to reduced expression of thermogenic genes, impaired β-adrenergic responsiveness, and compromised mitochondrial function, thereby limiting adipocyte metabolic flexibility [76,77]. Preliminary and recent studies indicate that GLP-1RAs reduce the abundance of pro-inflammatory M1 macrophages in VAT while promoting a shift toward anti-inflammatory M2-like macrophages. This immunological shift may lower pro-inflammatory cytokine levels that suppress key thermogenic regulators, including PGC-1α and PRDM16 [78-80]. By mitigating inflammatory signaling, GLP-1RAs may enhance adipocyte plasticity, promote mitochondrial biogenesis, improve oxidative metabolism and thermogenic capacity, and ultimately increase energy expenditure [81-83].
The immunomodulatory effects of GLP-1RAs appear to extend beyond macrophage polarization and may also involve other immune cell populations, including regulatory T (Treg) cells, eosinophils, and type 2 innate lymphoid cells (ILC2s) [84-86]. Preclinical studies suggest that these cells contribute to WAT browning and the formation of beige and brown adipocytes. Collectively, they provide a cellular framework that supports the development and maintenance of thermogenic AT through cytokine- and growth factor-mediated signaling pathways that regulate adipocyte differentiation and function. In addition to influencing individual immune cell populations, GLP-1RAs may reshape the overall immune composition of WAT, creating a microenvironment permissive for beige and brown adipocyte programming [87,88]. Accordingly, this immunometabolic reconfiguration of WAT is likely to operate in concert with neural and metabolic pathways governing thermogenesis, rather than as an independent process.
In summary, GLP-1RAs, as illustrated in Figure 2, reprogram WAT metabolism by attenuating local inflammation, restoring thermogenic signaling, and promoting the formation of beige and brown adipocytes. Through these effects, they enhance mitochondrial activity and energy expenditure, extending their benefits beyond the regulation of food intake and glycemic control.
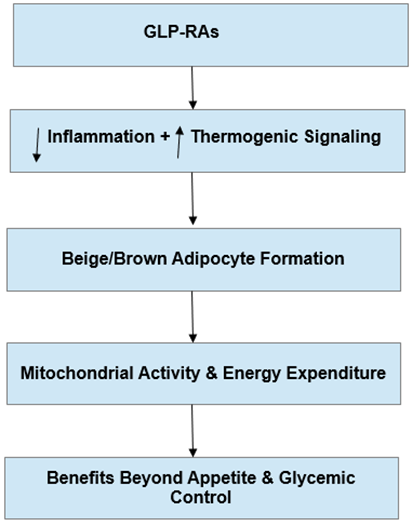
Figure 2: GLP-1 receptor agonist–mediated reprogramming of white adipose tissue metabolism. GLP-1 receptor agonists (GLP-1RAs) dampen local inflammation and restore thermogenic signaling, promoting beige and brown adipocyte formation and increasing mitochondrial activity and energy expenditure. Together, these effects extend metabolic benefits beyond appetite regulation and glycemic control. Upward arrows denote increases or activation, while downward arrows denote decreases or suppression.
Neurogenic Control of Sympathetic Activity by GLP-1 Receptor Agonists
One of the most researched mechanisms by which GLP-1 RAs influence AT remodeling is their effect on CNS signaling [89,90]. GLP-1Rs are concentrated in key hypothalamic and brainstem nuclei that regulate energy homeostasis, such as the paraventricular nucleus, arcuate nucleus, and nucleus of the solitary tract [91]. When stimulated by GLP-1RAs, these nuclei enhance sympathetic nervous system (SNS) activity directed toward peripheral tissues, including AT, increasing sympathetic nerve activity targeting fat deposits [92,93]. Sympathetic innervations significantly contribute to the formation of beige AT primarily through the activation of the β2-adrenergic receptor, which leads to cAMP production, PKA (Protein Kinase A) activation, and the initiation of several downstream transcriptional programs essential for thermogenesis [94,95]. Elevated SNS activity promotes increased lipolysis, mitochondrial biogenesis, and the expression of UCP-1 (Uncoupling Protein 1), all of which support the differentiation of beige adipocytes within WAT [96,97].
Therefore, CNS-induced sympathetic activation is likely a crucial regulatory pathway linking GLP-1 RA treatment with the thermogenic remodeling of AT. In this regard, Figure 3 on the following page illustrates CNS-mediated sympathetic activation by GLP-1 RAs, promoting β2-adrenergic signaling, thermogenic gene activation, beige adipocyte formation, and increased mitochondrial function in WAT.
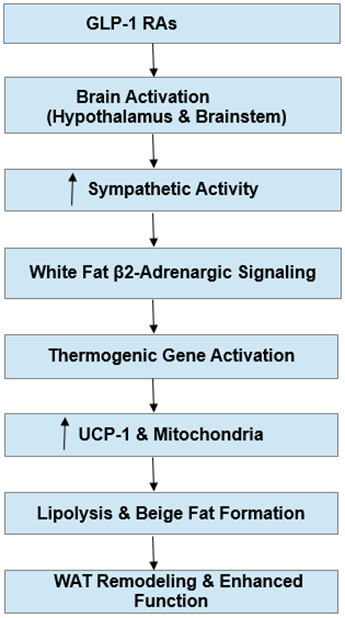
Figure 3: Central-peripheral mechanism of GLP-1 receptor agonist-mediated white adipose tissue remodeling. GLP-1 receptor agonists (GLP-1 RAs) stimulate neural circuits in the hypothalamus and brainstem, enhancing sympathetic nervous system output. This increased sympathetic drive engages β2-adrenergic receptors in white adipocytes, promoting expression of thermogenic genes, upregulation of uncoupling protein-1 (UCP-1), and mitochondrial biogenesis. The coordinated effects drive lipolysis, beige adipocyte formation, and overall remodeling of white adipose tissue, improving its metabolic function. Interlinked vertical arrows indicate the sequential signaling and downstream effects.
Transcriptional Regulation of Brown Adipocyte Commitment and Thermogenesis
The differentiation of brown adipocytes from preadipocytes proceeds through a conserved transcriptional cascade that regulates adipocyte lineage specification, commitment to the adipogenic fate, and the acquisition of thermogenic capacity [98,99]. Peroxisome proliferator-activated receptor gamma (PPARγ) functions as the master regulator of adipogenesis in both white and brown adipocytes; however, lineage-specific transcription factors, co-regulators, and accessory proteins modulate PPARγ transcriptional outcome in brown adipocytes, enabling a shift in gene expression toward mitochondrial biogenesis and thermogenesis [100,101]. Importantly, commitment to the brown adipocyte lineage is initiated by PRDM16, which activates brown adipocyte–specific genes while repressing myogenic and white adipogenic transcriptional programs [102]. This lineage decision is subsequently reinforced at the chromatin level by enhancer-specific transcription factors. Early B-cell factor 2 (EBF2) activates brown adipocyte enhancers and facilitates the preferential recruitment of PPARγ to thermogenic regulatory elements, whereas nuclear factor I-A (NFIA) further fine-tunes enhancer activation and PPARγ genomic occupancy [103,104]. In this context, PPARγ binding is supported by chromatin environments programmed by enhancers (via recruitment of transcription factors, co-activators, and chromatin-modifying enzymes such as histone acetyltransferases) that promote oxidative phosphorylation and mitochondrial gene expression in BAT, while simultaneously limiting inflammatory transcriptional programs [105,106].
Beyond sequence-specific transcription factors, emerging evidence indicates that transcriptional scaffolding proteins play an important role in organizing brown fat gene regulons. Among these, LIM domain-binding protein 1 (LDB1) has been identified as a transcriptional co-regulator that functions as a dimerized scaffold, facilitating the assembly of multiprotein transcriptional complexes at target loci [107,108]. Studies employing a global LDB1 heterozygous mouse model have uncovered previously unrecognized roles for LDB1 in the transcriptional regulation of BAT, suggesting a functional contribution to BAT biology [109,110]. However, direct molecular and functional evidence linking LDB1 to BAT thermogenesis and substrate utilization remains limited, raising the possibility that LDB1 supports BAT metabolism through transcriptional coordination. Furthermore, following lineage specification and enhancer programming, adipogenic commitment is driven by the early induction of CCAAT/enhancer-binding protein beta (C/EBPβ) and CCAAT/enhancer-binding protein delta (C/EBPδ), which remodel chromatin and induce PPARγ expression [111,112]. This phase is accompanied by sustained activity of PRDM16, EBF2, NFIA, and potentially transcriptional scaffolding proteins such as LDB1, culminating in the activation of brown adipocyte–specific gene expression [113,114]. Terminal differentiation is consolidated by CCAAT/enhancer-binding protein alpha (C/EBPα), which stabilizes the differentiated state through positive feedback with PPARγ [115,116].
Additionally, thermogenic differentiation requires the transcriptional coactivator PGC-1α, which enhances mitochondrial biogenesis and oxidative phosphorylation (OXPHOS) by coactivating PPARγ, nuclear respiratory factors, and estrogen-related receptors [117,118]. This program is further reinforced by PRDM16, which stabilizes PGC-1α at thermogenic regulatory elements, forming a feed-forward loop that sustains high mitochondrial content and uncoupled respiration [119,120]. To maintain appropriate control over this process, transcriptional repressors such as Forkhead box protein P1 (FOXP1) act to restrain thermogenic activation by antagonizing PPARγ- and PRDM16-dependent transcription [121,122].
Collectively, PPARγ, PRDM16, PGC-1α, C/EBP family members, EBF2, NFIA, FOXP1, and transcriptional co-regulators such as LDB1 constitute an enhancer-driven regulatory network that governs brown adipocyte differentiation, function, and metabolic activity. Through coordinated transcriptional organization, this network enables BAT to dynamically respond to environmental and metabolic cues that regulate systemic energy balance. All being said, the transcriptional hierarchy of brown adipocyte thermogenesis is well established, yet systemic metabolic signals such as GLP-1RAs may further fine-tune this network by modulating mitochondrial, inflammatory, and substrate-utilization pathways, as discussed in the following subsection.
GLP-1 Receptor Agonists as Modulators of Brown Adipocyte Transcriptional Programs
Data from in vitro models demonstrate that GLP-1RAs can directly modulate adipocyte transcriptional programs controlling differentiation and metabolic maturation [123,124]. In mesenchymal stem cell models such as C3H10T1/2, the GLP-1RA liraglutide promotes adipogenic, and particularly brown adipogenic, commitment, as indicated by increased cell proliferation, multilocular lipid accumulation, and induction of core adipogenic transcription factors such as PPARγ and C/EBPα, along with downstream adipocyte markers including FABP4/aP2 (Fatty Acid-Binding Protein 4 / adipocyte Protein 2) [125,126]. In parallel, increased mtDNA content and induction of mitochondrial and oxidative genes associated with brown adipocyte identity, including PGC-1α and UCP1, support a role for liraglutide in enhancing mitochondrial biogenesis and metabolic gene expression [127,128]. These effects are mediated through activation of the PI3K-AKT-mTOR (Phosphoinositide 3-kinase (PI3K)-Protein kinase B (AKT)-mechanistic target of rapamycin (mTOR)) signaling axis, as liraglutide induces phosphorylation of AKT and mTOR, key integrators of growth factor and nutrient signaling [129,130]. A causal link between GLP-1-responsive intracellular signaling and transcriptional regulation of adipocyte differentiation is further supported by pharmacological inhibition of PI3K with LY294002, which blocks downstream signaling and abrogates liraglutide-induced adipogenic differentiation, mitochondrial gene expression, and mtDNA expansion [131,132]. Consistent with these findings, studies in 3T3-L1 preadipocytes show that GLP-1 signaling enhances early adipogenic transcriptional cascades, including induction of PPARγ and C/EBPα, accompanied by increased Akt phosphorylation [133,134]. Together, these data support the concept that GLP-1-dependent signaling interfaces with canonical adipogenic transcriptional programs at early stages of differentiation, reinforcing lineage commitment rather than driving alternative cell fate decisions.
In contrast, in mature adipocytes, GLP-1RAs primarily modulate transcriptional states linked to metabolic function rather than lineage identity. Studies in human adipocytes and AT explants report reduced expression of adipogenic and lipogenic genes, alongside increased expression of genes involved in lipid turnover and insulin sensitivity following treatment with GLP-1 or GLP-1Ras [135,136]. These changes are often accompanied by suppression of inflammatory gene expression, suggesting that GLP-1RAs may alleviate inflammatory repression of PPARγ-dependent metabolic gene programs [137,138]. With respect to brown and beige adipocytes, current evidence supports an indirect mechanism of transcriptional regulation. Activation of the GLP-1R has been shown to enhance expression of thermogenic genes such as PGC-1α and UCP1 in BAT through sympathetic nervous system-dependent β-adrenergic signaling [139,140]. However, the extent to which GLP-1RAs directly activate thermogenic transcriptional programs in adipocytes remains incompletely defined. Importantly, GLP-1RA–induced transcriptional modulation appears to preserve the endogenous regulatory hierarchy governing adipocyte lineage identity. Rather than overriding lineage-specifying networks, GLP-1RAs act as transcriptional modulators, reinforcing mitochondrial, metabolic, and anti-inflammatory gene programs within an established PPARγ-centered framework [141,142]. Transcriptional regulators such as FOXP1 likely limit excessive thermogenic gene activation, maintaining transcriptional balance [143,144].
Together, these findings highlight the role of GLP-1RAs in fine-tuning adipocyte transcriptional states according to differentiation status and metabolic function, while underscoring the need to further distinguish direct versus indirect mechanisms of transcriptional control in brown and beige adipocytes.
Systemic Drivers of GLP-1 Receptor Agonist-Induced Adipose Tissue Remodeling
GLP-1RAs Preferentially Limit Adipocyte Hypertrophy Over Hyperplasia
Mounting evidence suggests that GLP-1 RAs, such as semaglutide and liraglutide, primarily regulate AT expansion by suppressing adipocyte hypertrophy rather than altering adipocyte hyperplasia [145,146]. AT growth occurs through two coordinated processes: enlargement of existing adipocytes (hypertrophy) to store excess lipid and the generation of new adipocytes from precursor cells, such as preadipocytes, through hyperplasia [147]. While hyperplasia supports healthy lipid storage, uncontrolled adipocyte hypertrophy promotes inflammatory and dysfunctional AT, a process that often emerges with aging due to impaired adipogenesis [148,149].
GLP-1RAs Enhance Adipose Tissue Integrity and Function
Consistent with this framework, several studies report that GLP-1RA treatment reduces adipocyte size, a phenotype typically associated with enhanced insulin sensitivity, and increases stromal support within AT [150,151]. Reductions in adipocyte volume and improvements in AT structural integrity correlate strongly with enhanced systemic insulin sensitivity and overall metabolic function [152,153]. Although some studies suggest that GLP-1RAs may influence adipocyte progenitor commitment or differentiation, accumulating evidence does not support a direct inhibitory effect on these processes, likely reflecting the low expression of GLP-1Rs in adipocytes [154,155].
GLP-1RAs Limit Fat Accumulation and Inflammation Indirectly
Mechanistically, GLP-1RAs appear to primarily limit the accumulation of free fatty acids (FFAs) within mature adipocytes [156,157]. Robust evidences for direct effects on adipocyte hyperplasia or adipogenesis remains lacking, with most studies reporting little to no change in total adipocyte number. These effects are thought to arise predominantly through indirect mechanisms, including reduced food intake, enhanced insulin action, and improved neuroendocrine signaling, which collectively restrict FFA influx and storage. By preserving the capacity for healthy AT remodeling while preferentially constraining excessive hypertrophy, GLP-1RAs prevent the transition of AT toward a proinflammatory state and confer metabolic benefits that extend beyond weight loss by alleviating lipid-driven inflammation and metabolic dysfunction [158,159].
GLP-1RAs Shape Adipocyte Lipid Metabolism
GLP-1RAs regulate AT structural remodeling, as reflected in functional transcriptomic changes in lipid metabolism. Experimental studies suggest interactions between GLP-1 RAs and key genes regulating fatty acid uptake (e.g., CD36, [FABP4/FABP5]) and de novo lipid synthesis (e.g., fatty acid synthase [FASN], stearoyl-CoA desaturase 1 [SCD1]), consistent with reduced lipid accumulation [160,161]. However, upstream lipogenic regulators such as sterol regulatory element-binding protein-1c [SREBF1/SREBP-1c] and acetyl-CoA carboxylase [ACACA/ACACB] show inconsistent modulation across studies. Similarly, some in vitro and animal studies report increased markers of triglyceride hydrolysis and fatty acid mobilization, including adipose triglyceride lipase [PNPLA2/ATGL] and hormone-sensitive lipase [LIPE/HSL] [162,163]. These effects, however, are not consistently observed in human AT, raising questions about the direct relevance of animal findings.
GLP-1RAs Boost Adipocyte Mitochondria and Promote Beiging
GLP-1RAs are linked to augmented oxidative and β-oxidation capacities in adipocyte mitochondria, reflected by upregulation of genes involved in OXPHOS and mitochondrial biogenesis, including peroxisome proliferator-activated receptor gamma coactivator 1-alpha [PPARGC1A/PGC-1α], PPARα, carnitine palmitoyltransferase 1A/1B [CPT1A/CPT1B], and acyl-CoA oxidase 1 [ACOX1] [164,165]. In certain adipose depots, partial beiging has been observed in animal models, accompanied by increased expression of PRDM16, UCP1, and cell death-inducing DFFA-like effector A [CIDEA], although evidence in humans remains limited [166,167]. Most of these gene expression changes are likely secondary to systemic effects such as reduced caloric intake, improved glucose handling, and altered neuroendocrine signaling, rather than direct activation of adipocyte GLP-1Rs.
Molecular and Metabolic Impacts of GLP-1 RAs Across Organs
GLP-1RAs affect not only AT but also limit lipid accumulation in organs such as the liver, skeletal muscle, and heart [168]. Their mechanism includes modulating expression of genes regulating lipid uptake (CD36, SLC27A/FATP1-6), lipid synthesis (FASN, SCD1), triglyceride storage (DGAT2), and lipotoxic lipid production (CERS2/CERS6), along with enhanced insulin signaling (IRS1/2, AKT2) and adiponectin (ADIPOQ)-mediated promotion of fatty acid oxidation [169,170]. Collectively, these molecular changes-mostly observed in preclinical and cellular studies-likely reflect systemic responses rather than direct activation of GLP-1Rs in adipocytes. Nevertheless, they align well with clinical observations, including improved lipid turnover, reduced circulating FFAs, decreased ectopic lipid deposition, and enhanced insulin sensitivity in patients treated with GLP-1Ras [171].
Neuroendocrine Control of Adipose Tissue by GLP-1RAs
A key component of the indirect effects of GLP-1RAs is central regulation of appetite and energy balance. GLP-1 RAs activate GLP-1Rs in hypothalamic and brainstem nuclei, which are abundant in these regions but sparse in AT. This activation engages anorexigenic pathways and suppresses orexigenic signaling, producing a chronic negative energy balance [172,173]. The resulting decrease in calorie intake gradually reduces adipose depot size and adipocyte volume. Smaller adipocytes exhibit higher insulin sensitivity and lipid-buffering capacity and are less prone to hypoxia-induced stress, restoring AT metabolic flexibility [174,175].
Improved Glucose Homeostasis Supports Adipose Metabolic Function
In parallel, GLP-1RAs enhance systemic insulin signaling and glucose regulation, supporting AT metabolic function. By increasing glucose-dependent insulin secretion and suppressing excess glucagon release, these drugs reduce glucotoxic stress and normalize insulin levels, improving cellular responsiveness [176,177]. Improvements in gut–hormone crosstalk and broader neuroendocrine adaptations further assist AT in managing lipid overload and metabolic stress [178,179].
Attenuation of Pro-Inflammatory Signaling by GLP-1RAs
GLP-1RAs also reduce pro-inflammatory responses in VAT, a key driver of obesity-associated metabolic disorders. Hypertrophic adipocytes and lipid accumulation trigger chemokine secretion, macrophage recruitment, and pro-inflammatory polarization, impairing insulin signaling and mitochondrial function. GLP-1RAs mitigate these effects by reducing adipocyte size and metabolic stress, indirectly lowering chemokine production and macrophage infiltration [180,181]. Preclinical studies show suppression of NF-κB (Nuclear Factor kappa-light-chain-enhancer of activated B cells) and JNK (c-Jun N-terminal kinase) pathways, with reductions in pro-inflammatory cytokines, and emerging human data confirm parallel decreases in circulating and AT-associated inflammatory markers alongside improved metabolic outcomes [182,183].
GLP-1RAs Deliver Multifaceted Adipose and Cardiometabolic Benefits
Taken together, these findings indicate that the metabolic benefits of GLP-1RAs extend beyond weight loss, reflecting qualitative improvements in AT structure, metabolism, and immunological status. The effects of GLP-1 RAs are best understood as the combined result of neuroendocrine regulation, sustained energy deficit, enhanced insulin signaling, and reduced inflammation, rather than solely direct activation of GLP-1Rs on adipocytes. This perspective explains why the cardiometabolic benefits of GLP-1RAs often exceed those expected from weight loss alone. Within this framework, Table 2 provides an overview of the multisystem effects of GLP-1RAs in metabolic regulation [184-194].
Table 2: Multisystem Effects of GLP-1 Receptor Agonists in Metabolic Regulation.
| Domain | Key Effects of GLP-1 RAs | Underlying Mechanisms | Physiological Outcomes | References |
| Overall metabolic role | Act as metabolic regulators apart from appetite suppression | Neuroendocrine signaling, tissue remodeling | Improved glucose control, cardiometabolic health | [184] |
| Adipose tissue (AT) remodeling | Convert WAT from pro-inflammatory storage tissue to metabolically active WAT | Prevent adipocyte hypertrophy, increase healthy adipocytes, reduce lipotoxic stress | Improved insulin sensitivity, enhanced energy expenditure | [185] |
| Immune modulation in AT | Shift macrophages from M1 → M2 phenotype | Reprogrammed immune metabolism | Ameliorate local and systemic inflammation | [186] |
| Thermogenesis & mitochondria | Induce beige/brown adipocyte traits | Activation of PPARγ, PGC-1α, PRDM16, EBF2, NFIA | Increased mitochondrial biogenesis, OXPHOS, uncoupled respiration | [187] |
| Weight-independent effects | AT benefits occur even without major weight loss | AT-targeted metabolic reprogramming | Improved metabolic flexibility of AT | [188] |
| Central regulation | Effects mainly mediated by CNS pathways | Given the limited expression of GLP-1 receptors in AT, the effects are likely indirect rather than direct. | Coordinated systemic metabolic control | [189] |
| Ectopic lipid reduction | Decrease fat accumulation in liver, muscle, hear | Improved lipid absorption, oxidation, adiponectin signaling, and reduced systemic inflammation | Reduced lipotoxicity, better insulin sensitivity | [190] |
| Gut microbiome | Increase beneficial taxa (Bifidobacterium, Lactobacillus) | Enhanced SCFA production, improved gut barrier | Reduced endotoxemia, lower inflammation | [191] |
| Gut–brain–adipose axis | Positive feedback loop via GLP-1 secretion | Microbial metabolites stimulate enteroendocrine cells | Sustained metabolic benefits | [192] |
| Limitations & gaps | Evidence largely correlative | Lack of mechanistic & longitudinal studies to establish causal relationship | Unclear role in sarcopenia prevention | [193] |
| Clinical implication | Systems-based precision therapy | Multi-organ pathway integration | Beyond weight loss: holistic cardiometabolic treatment | [194] |
Abbreviations: GLP-1 RAs, glucagon-like peptide-1 receptor agonists; T2D, type 2 diabetes; AT, adipose tissue; WAT, white adipose tissue; CNS, central nervous system; M1 macrophages, classically activated pro-inflammatory macrophages; M2 macrophages, alternatively activated anti-inflammatory macrophages; PPARγ, peroxisome proliferator-activated receptor gamma; PGC-1α, PPARγ coactivator 1-alpha; PRDM16, PR domain containing 16; EBF2, early B-cell factor 2; NFIA, nuclear factor I A; OXPHOS, oxidative phosphorylation; SCFAs, short-chain fatty acids.
Significance of The Review
GLP-1RAs extend beyond appetite and glucose regulation, promoting overall metabolic health by transforming AT into a more favorable, functional state. Through a combination of central and peripheral effects, these agents reduce visceral fat, enhance nutrient utilization, improve insulin sensitivity, and dampen inflammation, collectively supporting cardiometabolic well-being. This perspective suggests that focusing solely on weight loss may be too narrow, particularly in light of the obesity paradox, where higher body weight can confer survival advantages in certain populations. By redistributing AT to metabolically advantageous locations, health outcomes can improve even without substantial weight reduction. Individuals with poor metabolic health despite a low or average body weight-sometimes referred to as having “lean obesity”-may particularly benefit from GLP-1RA-induced changes in fat distribution, although direct clinical data in underweight populations remain limited.
Targeting the structure and function of AT opens new possibilities for incretin-based therapies that preferentially reduce visceral fat, preserve subcutaneous fat, and optimize adipokine profiles. By prioritizing metabolic improvements over mere weight loss, such approaches may also help reduce stigma around body size and foster a more inclusive view of health. This synthesis proposes a conceptual framework that recasts GLP-1RAs as AT-centered immunometabolic modulators rather than simply appetite suppressants or weight-loss agents. Immune reprogramming of WAT appears central to their cardiometabolic benefits, framing AT remodeling as an active therapeutic strategy rather than a passive consequence of weight change. By decoupling metabolic health from total body weight, this tissue-focused perspective challenges conventional weight-centric paradigms, encouraging clinicians to see GLP-1 RAs as tools for improving fat quality and distribution rather than solely reducing mass. Metabolic function, rather than body size alone, becomes the priority, and excess adiposity may not be inherently harmful if fat distribution supports healthy metabolic activity. Recognizing this could also help shift cultural and social perceptions around body ideals, especially in settings where higher body fat is traditionally associated with health.
Importantly, lower-dose GLP-1RA regimens that emphasize AT remodeling rather than maximal weight reduction may offer a cost-effective, sustainable option for long-term metabolic management. Such strategies could reduce risks linked to higher doses, including muscle loss, and help counter “obeseogenic memory,” the tendency for weight regain after therapy cessation [195,196]. When combined with healthy lifestyle habits, GLP-1RAs can improve AT function, reduce systemic inflammation, and enhance insulin sensitivity, independent of total weight change. Public health initiatives can leverage this understanding to promote interventions that improve fat tissue function, cardiometabolic resilience, and disease resistance—even among individuals not classified as overweight or obese.
Limitations of The Review
This review has several significant limitations. First, most of the existing knowledge about GLP-1RAs and their impact on AT remodeling is derived from preclinical studies conducted in rodents, where fat distribution, browning potential, and thermogenic activities vary from those in humans. Thus, the direct application of these findings to human physiology should be approached with caution. Second, although GLP-1RAs significantly decrease visceral adipose tissue and enhance metabolic parameters in humans, there is limited direct evidence supporting the browning of WAT or the activation of beige adipocytes. Most human data rely on imaging techniques, gene expression analysis, or circulating biomarkers instead of direct evaluations of thermogenic function. Additionally, there is a scarcity of clinical research involving individuals with normal or low body weight who still exhibit metabolic disturbances-often referred to as "lean obesity"-which complicates the ability to make definitive statements regarding the effectiveness and safety of GLP-1RA therapy in these groups [197,198]. Moreover, this review emphasizes mechanisms centered around AT and does not adequately consider inter-organ communication, long-term cardiovascular results, or real-world adherence and tolerance, all of which may affect the clinical outcomes of GLP-1RAs. Ultimately, the differences in study design, length of treatment, dosing schedules, and patient demographics in both preclinical and clinical investigations introduce additional complexity and may influence the understanding of the outcomes reported. In spite of these challenges, this review provides an extensive framework that identifies AT remodeling as a vital factor in the cardiometabolic advantages associated with GLP-1RAs and emphasizes important areas for future research.
Future Directions
GLP-1RAs clearly influence AT through a combination of central and peripheral mechanisms, yet the exact balance between direct effects on adipocytes and indirect systemic, neuroendocrine, or immune-mediated pathways remains unclear. To untangle these contributions, future studies should leverage tools such as cell-specific receptor knockout models, organoid systems, and human adipose explants. While preclinical studies in rodents provide strong evidence that GLP-1RAs promote adipose “browning” and upregulate thermogenic genes, direct evidence for the recruitment of beige adipocytes in humans is still limited. Confirmatory studies using human AT histology and molecular profiling are needed to translate these findings. Clinical data show that GLP-1RAs preferentially VAT and total fat mass in individuals with obesity or T2D, typically with only modest loss of lean mass. However, comparable data in normal-weight or lean individuals with poor metabolic health are scarce, highlighting the need for targeted trials in these populations. Long-term approaches using lower-dose or individualized GLP-1RA therapy may help balance metabolic benefits, safety, and cost while minimizing lean tissue loss. Research comparing different dosing regimens, combination therapies, and treatment durations will be important for developing precision medicine strategies. Translational and public health research should also incorporate AT biomarkers, advanced imaging techniques, and adipokine profiling to predict which individuals are most likely to benefit. This would allow interventions to focus on improving metabolic health rather than relying solely on BMI or weight-based metrics. Finally, given that GLP-1RAs influence metabolism in the liver, muscle, and cardiovascular system, future work should examine inter-organ communication and explore how coordinated transcriptional, immunometabolic, and neuroendocrine signaling drives systemic metabolic improvements [199,200].
Conclusion
GLP-1RAs are transforming our approach to metabolic treatment. Instead of merely reducing appetite and managing blood sugar levels, these medications actively alter adipose tissue and promote overall metabolic well-being. By decreasing visceral fat, improving nutrient utilization, enhancing insulin sensitivity, and lowering inflammation, GLP-1RAs can provide significant cardiometabolic advantages even in the absence of substantial weight loss. These benefits result from a complex interaction of immune system reprogramming, neuroendocrine communication, and gene expression regulation, emphasizing the importance of AT as a crucial therapeutic focus. Utilizing lower doses or tailored treatment strategies could provide lasting advantages while reducing side effects, highlighting a precision-focused approach that prioritizes metabolic function over just body weight. Moving forward, additional clinical and mechanistic studies across various populations will be crucial to fully unlock the capabilities of GLP-1RAs in enhancing overall metabolic health.
Declarations
Acknowledgments
The author would like to thank Dhurabjyoti Chattopadhyay for reviewing the work.
Funding
This research is supported by Bandhan, Kolkata, India.
Conflict of Interest
The author declares no conflict of interest.
References
-
-->























